Learning Objectives
- Place and identify land plants on a phylogenetic tree within the domain Eukarya
- Define and recognize traits and adaptations common to (nearly all) land plant taxa, including cuticle, stomata, roots/root-like structures, and mycorrhizal fungi
- Identify how key land plant adaptations including true roots, vascular tissue, lignin, pollen, seeds, and flowers are adaptations to increasingly drier environments
- Define, draw, and label the general alternation of generations life cycle
- Differentiate major land plant taxa (bryophytes, lycophytes, gymnosperms, and angiosperms) using the key adaptations to life on land and the dominant life cycle stage (gametophyte or sporophyte)
- Identify the geologic time periods when the major land plant taxa were dominant and why these periods are important to humans
Land Plants on the Phylogenetic Tree of Life
We have already spent quite a bit of time considering the evolutionary tree of life and the three domains of life, but we have focused mostly on domain-level evolutionary innovations that occurred in the Archaean and Proterozoic Eons. Now we will move into the Phanerozoic Eon (the current eon) and narrow in on one specific lineage of multicellular eukaryotes within the domain Eukarya: land plants.
Land plants are a monophyletic group of eukaryotic photoautotrophs. As eukaryotes, their cells contain a nucleus, mitochondria, and a complex system of internal membrane including the endoplasmic reticulum and Golgi apparatus; as photosynthetic eukaryotes, their cells also contain chloroplasts. The lineage that led to plants is thought to have diverged from the lineage that led to fungi and animals approximately 1.5 billion years ago during the Proterozoic Eon.
Note that we are specifically referring to LAND plants throughout this reading, such as mosses, ferns, conifers, and flowering plants. Algae, which are aquatic, photosynthetic eukaryotes, are also typically considered to be plants (though obviously not land plants); however, the term “algae” refers to a large and diverse group of photosynthetic eukaryotes that includes green, brown, and red algae that do not have a single common photosynthetic ancestor (in other words, the term “algae” is not monophyletic). But green algae and land plants do share a common photosynthetic ancestor: land plants evolved from a group of green algae approximately 480-470 MYA, which was relatively early in the Paleozoic Era within the Phanerozoic Eon. The common ancestry with green algae places plants on the phylogenetic tree of life as seen below:
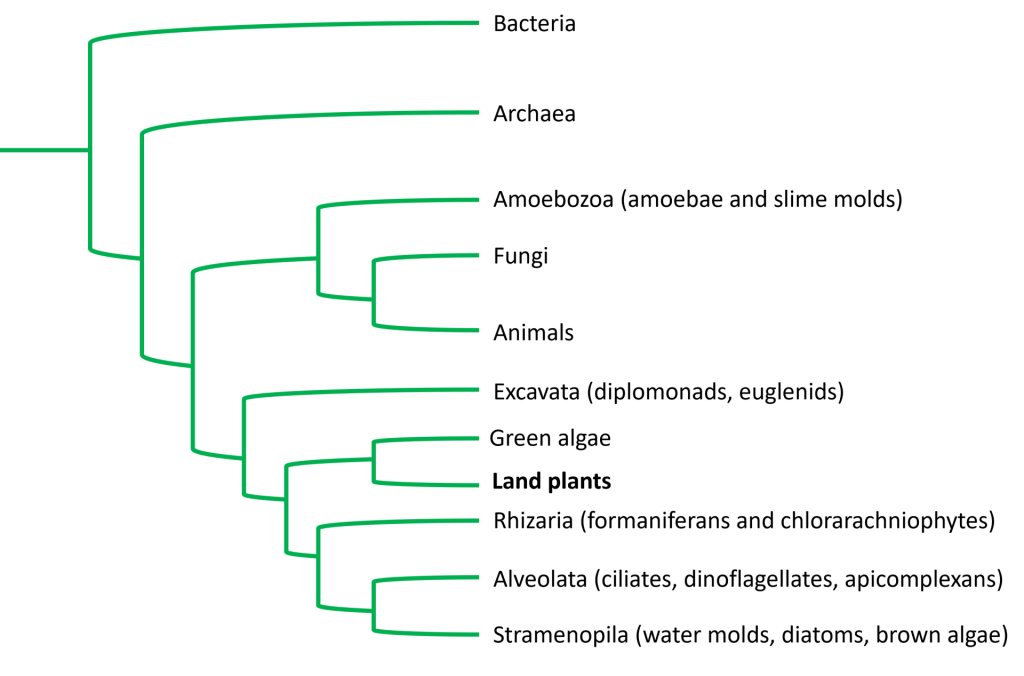
A more simplified tree of life, which does not show lineages of single-celled eukaryotes, would look like this:
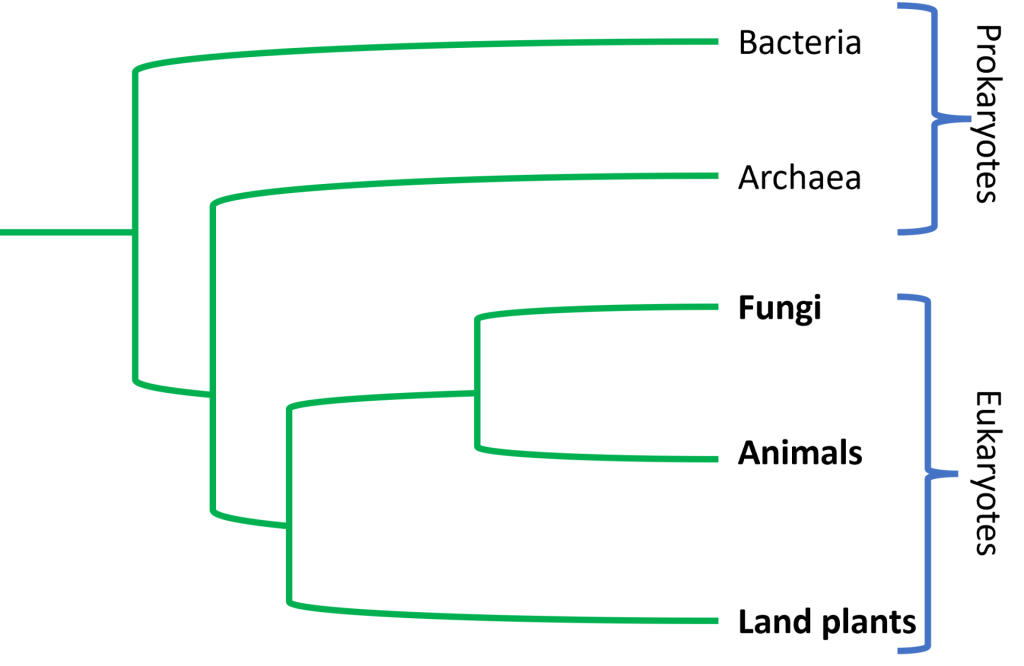
Challenges and Adaptations to Life on Land
The information below was adapted from OpenStax Biology 25.1
The ancestor of all land plants was an aquatic, green algal-like species with a cell wall composed of cellulose. Living in the water provides a number of advantages compared to life on land:
- In water or near it, plants can absorb water from their surroundings with no need for any special organ for absorbing the water, and no need for any special tissue to prevent desiccation (drying out).
- Water provides a sort of external support structure and buoyancy to living things; in contrast, living on land requires additional structural support to avoid falling over.
- Sperm and egg can easily swim to each other in a water environment, and do not need protection from desiccation. Sperm and egg require alternative strategies for a) getting to each other and b) avoiding drying out when on land.
- Water filters out a significant amount of ultraviolet-B (UVB) light, which is highly destructive to DNA (and one of the reasons to wear sunblock). No such filtering occurs in air, so terrestrial organisms require alternative strategies for protection against UV irradiation.
If life on land presents so many challenges, why did any land plants evolve to live on land? Life on land offers several advantages, including one particular advantage that was specific to the evolutionary history of life approximately 470 MYA early in the Paleozoic Era of the Phanerozoic Eon:
- Sunlight is abundant in air compared to water. Water acts as a filter, altering the spectral quality of light absorbed by the photosynthetic pigment chlorophyll.
- Carbon dioxide, the required carbon source for green plants, is more readily available in air than in water, since it diffuses faster in air.
- Land plants evolved before land animals; therefore, no predators threatened early plant life. This situation changed over time as animals colonized land, where they fed on the abundant sources of nutrients in the established flora. As a result of this selective pressure by plant-eating animals, plants evolved adaptations to deter predation, such as spines, thorns, and toxic chemicals.
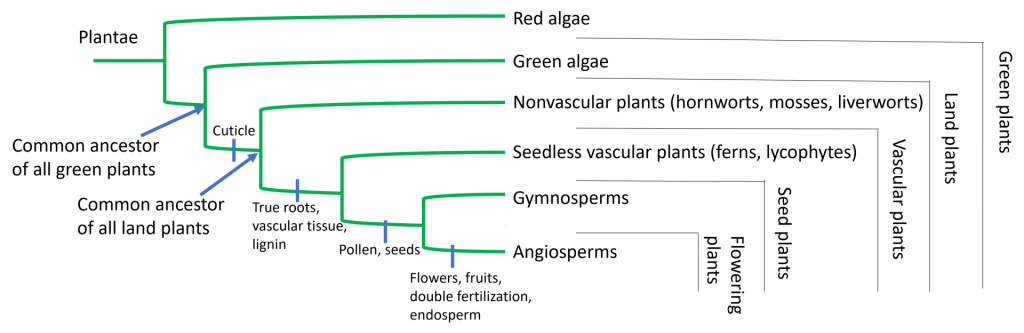
The evolution of specific adaptations in aquatic plants allowed them to transition to a terrestrial environment. In fact, modern land plants have an array of adaptations to life on land, but they did not evolve all at once. In addition, different adaptations are present in different plant lineages. The adaptations and characteristics which are universally present in (nearly) all land plants include:
- A waxy cuticle that covers the outer surface of the plant and prevents drying out through evaporation. The cuticle also partially protects against radiation damage from UV light. A waxy cuticle is universally present in all land plants, though it is much thinner in nonvascular plants called bryophytes (mosses, liverworts, and hornworts) compared to all other land plants.
- Stomata (singular: stoma) are pores or holes which allow for exchange of gasses (such as oxygen and carbon dioxide) between the plant cells and the environment. Stomata or similar structures are necessary in land plants because the waxy cuticle blocks free-flow of gasses. Stomata are present in all land plant lineages except liverworts (liverworts are similar to and closely related to mosses).
- Roots (or root-like structures) anchor plants to the soil and, in plants with true roots, serve as conduits for water absorption. All land plants except Bryophytes (mosses, liverworts, and hornworts) have true roots. Bryophytes have root-like structures called rhizoids that anchor them to their substrate, but are not involved in water absorption (their lack of true roots is one reason that Bryophytes can only survive in very moist environments).
- Mutualistic association with mycorrhizal fungi, which are tightly associated with plant roots. Mycorrhizal fungi are associated with approximately 80% of all land plant species and provide additional surface area for absorption of both water and nutrients from the soil. The fungi share these resources with the plant roots, and, in exchange, the plant shares photosynthetic sugar products with the fungi. In fact, the evolution of land plants is intertwined with the evolution of fungi on land in another way beyond root associations: fungi helped break down the rock on early land to create soil, which was required for plants to be able to begin growing on land.
- The alternation of generations life cycle, which includes both a multicellular haploid stage and a multicellular diploid stage. Why is this an adaptation to life on land? It isn’t, on its own, a true adaptation to life on land (in fact, alternation of generations also occurs in some green algae, which are aquatic but share a common ancestor with all land plants). But specific alterations to the steps of the alternation of generations life cycle have occurred in different lineages of plants, and those alterations DO function as adaptations to life on land. We’ll consider these adaptations later in this reading.
Key Adaptations to (Increasingly Drier) Life on Land
The information below was adapted from OpenStax Biology 25.1
Early land plants could not live very far from an abundant source of water. Over evolutionary time, land plants evolved adaptations to that allowed them to survive increasing degrees of dryness and grow farther from ready sources of water:
- Nonvascular plants, or Bryophytes (liverworts, mosses, and hornworts) are essentially physically tied to water. Their major adaptions to life on land include a waxy cuticle and root-like structures called rhizoids, but they lack true roots. Other than these two traits, they are heavily dependent on water for their life cycle:
- Nonvascular plants must live in very moist environments near sources of water.
- They are very short because they have no mechanism for transporting water up against gravity.
- Their sperm and eggs require water for mating: the gametes are not protected from desiccation, and the flagellated sperm must swim in water to find the egg.
- Seedless vascular plants (lycophytes, ferns, and horsetails) have three major adaptations compared to nonvascular plants: true roots, vascular tissue, and lignin. These adaptations allowed seedless vascular plants to outcompete nonvascular plants in early colonization of life on land.
- True roots grow deeper into the soil than rhizoids, allowing for better extraction of water and nutrients from the soil.
- Vascular tissue (xylem and phloem) consists of tube-like cells that allow for transport of water (in xylem) from roots to leaves and transport of sugars (in phloem) from leaves to the rest of the plant tissues. The adaptation of vascular tissue meant that these plants could grow taller than bryophytes (and thus get more access to sunlight for photosynthesis).
- Lignin, a rigid component of some plant cell walls that provides structural rigidity and allows for higher movement of water against gravity and thus taller plant growth, also first evolved in these groups.
- Aside from these three adaptations, seedless vascular plants are still tied to the water for reproduction: like nonvascular plants, their sperm and eggs are sensitive to desiccation and the sperm must swim through water to get to the egg.
- Seeded, nonflowering plants, or gymnosperms, (ginkgos, cycads, and conifers) are trees that grow to greater heights on land by combining the strength of lignin with the phenomenon of secondary growth (e.g. tree rings – getting wider). They have two additional adaptations beyond seedless vascular plants: pollen and seeds. These two adaptations allowed them to colonize drier habitats than nonvascular and seedless vascular plants:
- Pollen, a mechanism for delivering sperm to egg in the absence of water. Many sources state that pollen is the same thing as sperm, which is not true. In fact, pollen produce sperm. We’ll revisit this point later on in this course, so for now, just know that pollen protects sperm from desiccation and provides a means for sperm to reach the egg in the absence of water. Seeded nonflowering plants typically rely on wind to transport pollen (and therefore sperm) to eggs.
- Seeds, which protect the fertilized egg on land in multiple ways. Most obviously, the seed is a hard physical structure which protects the fertilized egg (embryo) against desiccation (drying out). But in addition, and less obviously, the seed is a form of ‘suspended animation’ for the embryo that pauses development until environmental conditions are favorable for seed germination, or emergence of the embryo from the seed to start growing as a plant.
- Flowering plants, or angiosperms, possess the most recent adaptations to life on land: the flower, double fertilization and the endosperm, and fruit:
- Flowers might not seem like an obvious adaptation to living on land, but flowers allow for pollinators (such as insects, birds, bats, and other animals) to move pollen (and therefore sperm) to eggs. Because reliance on pollinators is much less random than reliance on wind, dramatically increasing likelihood of successful pollination, flowers represent an important adaptation to life on land. In some instances, flowers have also driven co-evolution between different flowering plants species and their specific pollinators.
- Double fertilization and the endosperm: Double fertilization is a unique process in flowering plants, where one sperm fertilizes the egg to create an embryo, and a second sperm fertilizes another structure next to the egg to create an endosperm. The endosperm undergoes a sort of pseudo-development, where it increases in mass and contents to create a nutrient reserve which will later provide nutrients to the developing embryo during germination. We will discuss these concepts in much greater detail later in the course.
- Fruits are any structure that aid in seed dispersal, such as something sweet that is eaten by an animal so that the seed is deposited somewhere new in the feces (and with its own personal supply of fertilizer!). Fruits thus provide a mechanism for seeds to colonize new territories away from the parent plant.
The phylogenetic tree below shows the evolutionary relationships between modern plants, as well as the origins of adaptations in each plant lineage:
Adaptations within the Alternation of Generations Life Cycle in Land Plants
As we’ve previously discussed, all eukaryote sexual life cycles include a haploid stage and a diploid stage. Usually one of these stages is large and multicellular (the organism we can see by eye), while the other is small and unicellular. All land plants (and some green algae) reproduce via the alternation of generations life cycle, where both the haploid and the diploid stage of an organism are multicellular: the haploid multicellular form, called the gametophyte, is followed in the life cycle sequence by a multicellular diploid form, called the sporophyte. The sporophyte is named for the fact that it produces spores, which then grow into the gametophyte, which is named for the fact that it produces the gametes (egg and sperm cells).
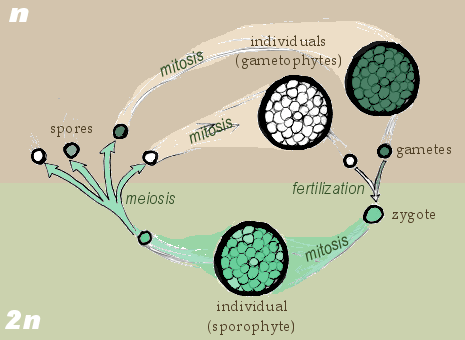
Though all plants display an alternation of generations life cycle, there are significant variations in different lineages of plants based on their evolutionary history. As you review the information below, make note of the specific adaptations that allowed colonization of increasingly dry environments over land plant evolutionary history:
- In seedless non-vascular plants, or bryophytes, the haploid gametophyte is larger than the sporophyte (the plant structure that you see is the gametophyte); this is a gametophyte-dominated life cycle. (By “dominated” we mean “the stage of the plant you can see by eye.”)
- The haploid gametophyte is adapted for photosynthesis, and is also the location of fertilization to generate the diploid sporophyte. The gametophyte then supports the sporophyte, which is attached to and dependent on the gametophyte for water and nutrients.
- The diploid sporophyte produces haploid spores, which are adapted for drying out and being dispersed to a new environment, where, if they land in a favorable location, will grow into a new gametophyte to start the cycle anew.
- In seedless vascular plants (such as ferns), the sporophyte is larger than the gametophyte (the plant structure that you see is the sporophyte), but the gametophyte is free-living and independent from the diploid sporophyte.
- The gametophyte in seedless vascular plants is very small, and it is the site of fertilization to generate the diploid sporophyte.
- The diploid sporophyte initially begins its development on the gametophyte; however, it grows larger and becomes photosynthetic; ultimately it produces haploid spores which are adapted for drying out and dispersal to a new environment.
- The life cycle of gymnosperms (conifers) and angiosperms (flowering plants) is dominated by the sporophyte stage (the plant structure that you see is the sporophyte), with the gametophyte remaining attached to and dependent on the sporophyte (complete reverse of bryophytes):
- The diploid sporophyte is large and photosynthetic; it produces haploid spores that grow into small haploid gametophytes.
- The female gametophyte remains associated with the sporophyte; the male gametophyte (pollen) is adapted for drying out and is carried to the female gametophyte by wind, water, or animal pollinators.
- The male and female gametophytes both produce gametes, which then fuse and develop into a diploid embryonic sporophyte within a seed; the seed is adapted for desiccation and dispersal to a new environment.
- Though they both have sporophyte-dominated life cycles, angiosperms and gymnosperms differ in that angiosperms have flowers, fruit-covered seeds, and double fertilization, while gymnosperms do not have flowers, have “naked” seeds, and do not have double fertilization.
The video below describes the features of nonvascular plants (mosses, liverworts, hornworts), and their alternation of generations life cycle:
And the video below describes the features of vascular plants and their alternation of generations life cycle.
(Note that the video below seems to suggest that seeded vascular plants produce seeds instead of spores, but this is incorrect! Seeded vascular plants do produce spores (just like all plants do); what is different is that the seeded plants use seeds for dispersal instead of using spores for dispersal; they still produce spores, but their spores remain associated with the sporophyte instead of dispersing to a new environment.)
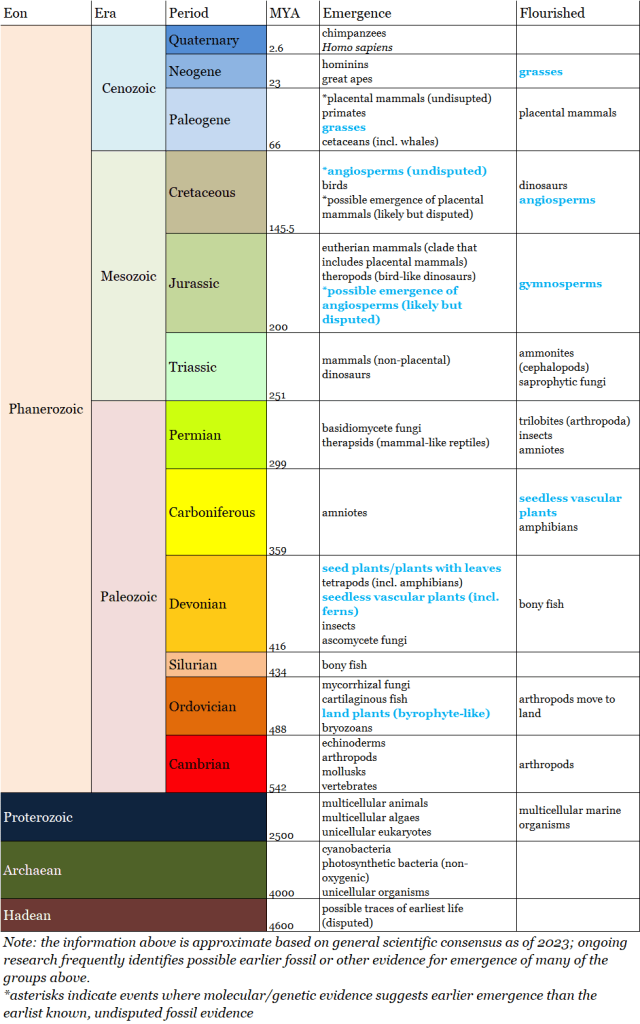
Plant Evolution over Geologic Time
The information below was adapted from OpenStax Biology 25.1
Before we discuss evolution of plant lineages over geologic time, first let’s briefly review the relevant eras and periods of the Phanerozoic. The Phanerozoic is the Eon we are currently in, and it is divided into three Eras: the Paleozoic (oldest era), the Mesozoic (middle era) and Cenozoic (most recent and current era).

- The early era of the Phanerozoic is called the Paleozoic, and it is divided into six periods: beginning with the Cambrian period, followed by the Ordovician, Silurian, Devonian, Carboniferous, and Permian.
- The middle era of the Phanerozoic is called the Mesozoic, and it is divided into three periods: beginning with the Triassic, followed by the Jurassic and the Cretaceous.
- The most recent and current era of the Phanerozoic is the Cenozoic, and it is divided into to three periods: beginning with the Paleogene, followed by the Neogene and ending with the Quaternary. (The Paleogene and the Neogene were previously referred to as the Tertiary, which is now considered and obsolete term.
Below we highlight some of the key events in land plant evolution; the periods denoted in bold have particular relevance to humans:
- Ordovician period (Paleozoic era): Fossil evidence of land plant evolution begins in the Ordovician, when fossils of the ancestors of modern plants first appear in the fossil record. These plants would have been nonvascular plants, lacking true leaves or roots and living in extremely damp environments close to water.
- Devonian period (Paleozoic era): The Devonian period saw the first vascular plants early in the period, and the first seed-bearing plants by the end of the period:
- The oldest-known vascular plants have been identified in deposits from the Devonian. These now-extinct vascular plants probably lacked true leaves and roots and formed low vegetation mats similar in size to modern-day mosses, although fossils indicate that some reached up to one meter in height.
- Gymnosperms, the earliest seed plants, also first appeared in the fossil record during the Devonian. However, seed plants were not very abundant at this time as seedless vascular plants had already colonized the land in the wet Devonian climate.
- Carboniferous period (Paleozoic era): Fossil evidence indicates that the Carboniferous Earth was covered in green: the land was dominated by trees and forests including both seedless and seed-bearing plants. Seedless vascular plants formed large swamp forests that covered most of the land, and were composed of tall trees reaching up to 100 ft. tall. The Carboniferous is named for the extensive coal deposits that resulted from these massive and abundant swamp forests. The video below describes the impact and legacy of vegetation during Carboniferous period:
- Permian period (Paleozoic era): The Permian period had much drier climates than the earlier periods of the Paleozoic, which allowed seed-bearing gymnosperms to outcompete seedless vascular plants which had previously dominated the land. Many plants became extinct during the end-Permian mass extinction event.
- Jurassic period (Mesozoic era): Gymnosperms reached their greatest diversity and abundance during the Jurassic; the Jurassic can be considered the age of gymnosperms as much it is the age of dinosaurs. There is also some recent fossil evidence that flowering plants (angiosperms) may have first appeared during the Jurassic, but the evidence is preliminary and this conclusion is disputed.
- Cretaceous period (Mesozoic era): Undisputed fossil evidence indicates that flowering plants existed during the early Cretaceous, and they had become dominant by the mid-Cretaceous. Many modern groups of insects also appear in great abundance during the Cretaceous, including those that serve as pollinators. The video below describes evolution of flowering plants:
- Cenozoic era (we will consider the Cenozoic at the level of the era (rather than delving into the periods of this era): Fossil evidence indicates that grasses evolved early in the Cenozoic era and rapidly expanded and dominated the landscape; evolution of grasses correlated with evolution of both large grazing mammals and large predator mammals.
Here is the summary of these significant events in plant evolutionary history (in blue) on our geologic time scale: