Learning Objectives
- Differentiate between internal and external fertilization and identify environmental and organismal traits associated with each type of fertilization
- Explain why females are more likely than males to be “choosy” when selecting a mating partner
- Define adaptation, biological fitness, sexual selection, and sexual dimorphism, and recognize that sexual selection can operate on any sexually-reproducing population
- Differentiate between direct male competition, indirect male competition, and female choice, and identify examples and advantages of each
- Define and recognize examples of parental investment
- Differentiate between animal mating systems and predict relationships between these mating systems and sexual dimorphism or sperm competition
Sexual Reproduction: Fertilization
Sexual reproduction starts with the combination of a sperm and an egg in a process called fertilization. Fertilization can occur either inside (internal fertilization) or outside (external fertilization) the body of the female.
External Fertilization
The information below is adapted from OpenStax Biology 43.2
External fertilization usually occurs in aquatic environments where both eggs and sperm are released into the water, a process called spawning. Water protects the eggs from drying out during development. Species that utilize external fertilization might be fully aquatic, like fish and many aquatic invertebrates, or they may be partially aquatic and partially terrestrial (land-based), like many amphibians. For both types of organisms, getting the sperm and egg together requires that the gametes be released at the same time and in the same location to increase the likelihood of fertilization (otherwise the time and energy to produce all those gametes was wasted). How does that happen?
- In some species, including some fish, crustaceans, mollusks, and other invertebrates, there are environmental (water temperature, length of daylight) or biological (pheromones) cues that cause males and females to release gametes at the same time. In this situation, males and females are often not interacting with each other as individuals, but massed together so that all sperm and all eggs are in the same location.
- In other species, including many amphibians, individual males court individual females to induce the female to release the eggs, at which point the male releases the sperm to fertilize that individual female’s eggs.


Internal Fertilization
The information below is adapted from OpenStax Biology 43.2
Internal fertilization occurs most often in terrestrial animals, although some aquatic animals like aquatic mammals (whales, dolphins) and some fish (including some cartilaginous fish like sharks, and some bony fish like guppies) also reproduce via internal fertilization. There are three ways that offspring are produced following internal fertilization:
- Fertilized eggs are laid outside the female’s body and develop there, and the embryo receives nourishment from the yolk that is a part of the egg. This process occurs in most bony fish, many reptiles, some cartilaginous fish, most amphibians, two mammals, and all birds.
- Fertilized eggs are retained inside the female’s body, and the embryo receives nourishment from the egg’s yolk. The young are fully developed when they ‘hatch’ within the female’s body and then exit to the environment. This process occurs in some bony fish, some sharks, some lizards, some snakes, some vipers, and some invertebrate animals.
- Fertilized eggs are retained inside the female, and the embryo receives nourishment from the mother’s blood through a placenta. The offspring develops within the female’s body and may or may not be fully developed prior to birth, depending on the species. This process occurs in most mammals, some cartilaginous fish, and a few reptiles.
Internal fertilization has the advantage of protecting the fertilized egg from dehydration on land. Depending on the species, the embryo may also remain within the female, which protects the eggs from predation. Internal fertilization also increases the likelihood of fertilization by a specific male. Fewer offspring are produced through this method, but their survival rate is likely to be higher than that for external fertilization.
Females are “Choosy;” Males are Not
The statement above is a gross generalization, but biologically it tends to be true across most species that reproduce sexually, whether they reproduce with internal or external fertilization. Why is this the case? Generally females invest more in the offspring than a male does, and she has a limited number of eggs compared to practically limitless sperm produced by a male. If a female mates with a male of poor genetic quality, and if her offspring don’t survive as a result, then she has wasted a lot of energy and resources and ended up with nothing; evolutionarily, females in this situation are unlikely to pass on their traits to many offspring. But a male can mate with practically unlimited numbers of females with little loss of energy or resources, regardless of how successful those offspring may be. In other words, eggs are “expensive” and sperm are “cheap.” Thus, generally a female maximizes her reproductive success by mating with the “best” male she can, while generally a male maximizes his reproductive success by mating with as many females as possible. In other words, evolution typically selects for “choosiness” in females but not necessarily in males.
Any situation where one sex (or both sexes) select specific individuals to mate with (compared to randomly mating with any individual) will result in a phenomenon called sexual selection. Sexual selection is a type of natural selection where one sex has a preference for certain characteristics in individuals of the other sex, thus leading to increased reproductive success of individuals who have that particular characteristic.
It is important to keep in mind that adaptations (anything that increases an individual’s reproductive success) occur without conscious thought or intention on the part of the individual; see the Biological Principles textbook pages on “What is Evolution?” and “Evolution by Natural Selection” for help with this often confusing concept. The result of these types of selection is the evolution of different strategies for maximizing biological fitness. Biological fitness can be defined as reproductive success relative to others in the population. An individual who has, for example, 10 surviving offspring has higher fitness than an individual who has 7 offspring surviving offspring (even if the second individual started out with more than 10 offspring).
Males Compete for Access to Females
Because females of most sexually reproducing species are “choosy,” females are often the sex that sexually selects traits in males. As a result, males compete with each other for access to females and/or induce a specific female to mate with him. As a result of this competition, sexual selection often leads to sexual dimorphism, or distinct differences in size or appearance between males and females. These differences in size or appearance are called secondary sexual characteristics, exaggerated or showy, physical traits that are associated with mating behaviors and reproductive success. Examples include breasts, showy tails and headpieces, and crazier (to us) traits like the length of the eye-stalks in stalk-eyed flies.

Sexual dimorphism can also lead to specific behaviors in males that increase their reproductive success. In both cases, significant energy is spent in the process of locating, attracting, and mating with the sex partner.
Competition among males occurs whether species mate via internal or external fertilization. In species that mate via internal fertilization, it’s pretty obvious that multiple males can’t mate with a female at the same time, and thus they must compete with each other. In species that mate via external fertilization, the female controls how and when the eggs are released, and thus males must compete for access to her eggs outside of her body.
Direct Male Competition
Males often engage in direct male competition over potential mating partners. This type of competition occurs when females mate only with a single male, typically the “winner” of the competition. Direct male competition often includes aggression (fighting) between males, but there are other forms as well. Examples of direct male competition include:
- Male-male aggression: males fight with each other for access to females, like two male lions competing for access to a pride of females
- Courtship rituals: males engage in “dances” or other displays to attract females, a male bower bird building an elaborate nest to attract a female
- Lekking: a specialized form of courtship ritual where many males gather together in one place and “display” at the same time, allowing females to choose among them, like male peacocks gathering together and displaying their tails to females

Ken Clifton/Flickr
It’s not all just competition between males; females choose which males to mate with based on observing the male competition. Selection of the “best” male by females is called female choice or intersexual selection. Female choice (intersexual selection) and direct male competition (intrasexual selection) usually lead to selection for extremely “showy” traits that don’t appear to provide any benefit to the individual’s survival, and might even make it more likely for the animal to be eaten by a predator (think of the peacock’s tail – see below). But if the trait improves the male’s ability to produce successful offspring because more females choose to mate with him, then these traits do in fact improve an individual’s biological fitness, even at the cost of decreasing its survival!
One question is why females should “care” about these showy male traits. A leading hypothesis to answer this question is the good genes hypothesis, which is the idea that these sexually-selected, showy male traits are “honest indicators” of good genetic quality. In other words, it takes good genes to make a big flashy tail (and to avoid being eaten by a predator, since that big tail slows him down), so the bigger and showier the tail, the “better” the male.

This video gives a brief overview of the implications of the good genes hypothesis and sexual selection in humans:
Indirect Male Competition
Instead of (or in addition to) competing directly with each other to have the opportunity to mate with a female, males can also compete for fertilization of a female’s eggs after mating has already occurred. Competing after mating is also called indirect male competition, or sperm competition, and it results in one male being more successful than another at fertilizing a female’s eggs. This type of competition occurs in species where the female is likely to mate with multiple males, so instead of males directly competing with each other, they are competing via their sperm. In other words, if a female mates with more than one male, then any male whose sperm end up fertilizing more eggs is going to have more offspring, on average, than other males.. So if there is a trait that makes this male’s sperm more successful than other male’s sperm, then that trait is going to end up increasing in the population over generations.
Examples of traits and behaviors which typically confer first male advantage include:
- Mate guarding: a male remains close by the female after mating, preventing other males from mating with her until there has been time for his sperm to fertilize the eggs
- Copulatory plugs: the male’s ejaculate includes a sticky residue which temporarily blocks entry to the female’s reproductive tract, making it difficult for other males to mate with her until after there has been time for the first male’s sperm to fertilize the eggs
Examples of traits and behaviors which typically confer second male advantage include:
- Elaborate penis morphology: elaborate structures on the penis help remove the sperm of previous males from the female’s reproductive tract by essentially scraping out the previous ejaculate
- Large ejaculate volume and large testes: a large volume of ejaculate helps to flush out the sperm deposited in the female’s reproductive tract by the previous male; a large ejaculate volume means the testes must also be large, in order to have enough space to hold all the sperm

Female anatomy can also influence the success of sperm from specific males in a process called cryptic female choice, where a female is capable of preferentially using sperm from a specific male even if she has mated with multiple males. This process is poorly understood but suggests that males competition alone does not dictate success of that male sperm in fertilizing an egg.
This video provides a great overview of sperm competition, but be aware that it erroneously refers to bonobos as having a polygymous mating system (they are promiscuous) and gorillas as being monogamous (they are polygynous):
Parental Investment
Parental investment is any energy, effort, or resource that a parent provides to increase the offspring’s chances of survival, but at the cost of the parent’s ability to invest in other offspring. For example, a male songbird will collect food to feed his hatchlings, but he can only collect so much food in a given day. His investment of that food in one nest of hatchlings means that he cannot support hatchlings in a second nest at the same time; in this scenario, he supports one nest at the expense of another. Parental investment can include all types of parental care, as well as energy resources deposited in the egg or other nutrition provided to the developing embryo. It occurs both in species that reproduce via internal fertilization as well as those that reproduce via external fertilization. Some examples are shown below:


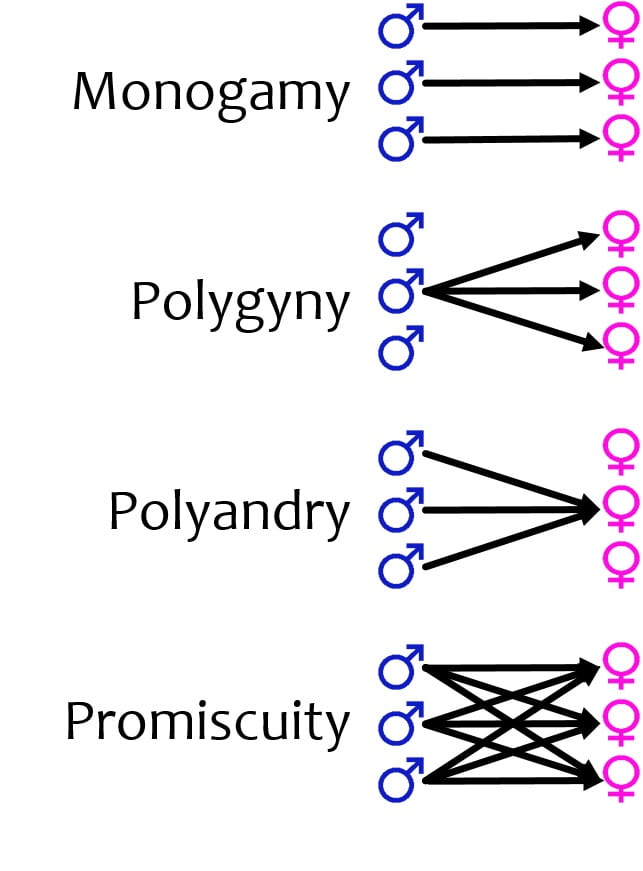
Animal mating systems
Sexually reproducing animals can be categorized by mating system, which is a general way of describing how males and females pair when mating. Mating systems are influenced by evolutionary history and environmental/ecological factors, and they in turn influence selection for different types of traits and behaviors in the species. There are three general mating systems, all involving innate and evolutionarily selected (as opposed to learned) behaviors, seen in animal populations: monogamous, polygamous, and promiscuous. The polygamous system includes two sub-types: polgynous and polyandrous systems.
Monogamy: In monogamous systems, one male and one female are paired for at least one breeding season. In some animals, such as the prairie vole, these associations can last beyond one breeding season and even as long as the breeding pair’s lifetimes. While there are many non-mutually exclusive hypotheses to explain selection for monogamous mating systems, one prominent explanation is the “male-assistance hypothesis,” where males that remain with a female to help guard and rear their young will have more and healthier offspring. The male-assistance hypothesis is supported by the observation that many monogamous species live in environments with widely scattered resources, meaning that it takes the effort of more than one adult to forage for enough resources to rear the young.

True monogamy, also called sexual monogamy, is where both partners mate only with each other. Assuming a 50/50 ratio of females to males, true monogamy is the only mating system where there is likely to be no competition for mates. True monogamy is exceedingly rare in biological systems. Prairie voles are one of the species that appear to exhibit a true monogamy mating system.
Social monogamy, where two individuals partner together to rear their offspring but also engage in “extra-pair copulations” (matings with other individuals,) is much more common that true monogamy. The vast majority of songbirds demonstrate social monogamy, where up to 40% of the offspring in a mating pair’s nest were not actually fathered by the male partner. Although each individual is, in principle, guaranteed a mate in social monogamy, there is still some amount of competition for mates in this mating system.
Social monogamy has both advantages and disadvantages for each partner, and these advantages and disadvantages are often related to competition for mates:
- You can imagine the advantage for a male in this scenario: he helps rear offspring with his social partner, increasing the likely survival of those offspring; but he also mates with other females, thus increasing his total number of offspring (assuming any of these other offspring also survive).
- Social monogamy can also be advantageous for the female: she has help from a social partner in raising her offspring, but she can also mate with other males who may be genetically “better.”
- The disadvantage for the male in this scenario is that he is most likely helping to raise offspring that are not his own.
- The disadvantage for the female is that the male may abandon her – and her offspring – if he detects that she has mated with another male.

Polygamy: Polygamy refers to either one male mating with multiple females or one female mates with many males, and includes the subcategories of polygyny and polyandry.
Polygyny: When one male mating with multiple females, called polygyny (“many females”), the female takes responsibility for most of the parental care as the single male is not capable of providing care to that many offspring. For instance, imagine that a male has established a territory such that he can provide access to resources. Females that enter the territory are drawn to its resource richness, which may signal that he has good genes for protecting a territory. The female benefits by mating with a genetically fit male at the cost of having no male help care for the offspring. For example, in the yellow-rumped honeyguide (a bird) males defend beehives because the females feed on beewax. As the females approach to find beeswax, the male defending the nest will mate with them.
Harem mating structures are a type of polygynous system where certain males dominate mating while controlling a territory with resources. In elephant seals, the alpha male dominates the mating within the group. Another type of polygyny is a lek system. In leks, the species has a communal courting area where several males perform elaborate displays for females, and the females choose their mate from the performing males. Lekking behavior is observed in several bird species including the sage grouse and the prairie chicken.

Polyandry: The other type of polygamy is called a polyandry (“many males”), where one female mates with multiple males. Polyandry very rare in biological systems, because it involves sex role reversal where females invest less in offspring while males invest more. Pipefishes, a close relative of seahorses, are an example of a species that exhibits polyandry where females compete for access to males. In both pipefishes and seahorses, males receive the eggs from the female, fertilize them, protect them within a pouch, and give birth to the offspring (see below). However, seahorses are monogamous, while pipefish are polyandrous.
Why do these two similar species differ in mating system? Ecologically, seahorses live in habitats with widely distributed resources, which means that the seahorse population is spread out and spread thin. The scattered population means that it is can be difficult to find a mating partner. Natural selection favors keeping a partner, once found, for reproductive assistance. In contrast to seahorses, pipefish tends to live in very dense populations in resource-rich environments. Because the male’s pouches, rather than the female’s eggs, are the limiting resource in reproduction, females compete with each other for access to males.

Promiscuity: Promiscuous mating systems occur when each female mates with multiple males, and each male mates with multiple females. Promiscuity generally occurs when a single male is unable to sexually monopolize a group of females, either because the females range more widely than the territory size of a single male, so they interact with multiple males (eg, the maximum territory size a male can defend is smaller than the females’ ranges), or because males and females live together in large social groups that a single male cannot monopolize.
In large social groups, often all females are sexually receptive at the same time, meaning that a single male cannot prevent other males from mating with other females while he mates with one female. Because each female mates with multiple males, paternity is never certain. The uncertainty of not knowing “who’s the daddy” selects for males to avoid infanticide, as they may inadvertently kill their own offspring.

CC BY-SA 4.0, https://en.wikipedia.org/w/index.php?curid=50736326
The video below provides a quick overview of animal mating systems:
Mating systems are influenced by competition for mates, and competition for mates is influenced by mating system. Except in the case of sexual (true) monogamy, there is always competition for fertilization. What differs in different mating systems is whether the competition occurs before mating (direct male competition) or after mating (sperm competition). In class we’ll spend some time considering the relationships between mating system, when competition occurs, and the resulting effects on an individual’s behavior and/or appearance.