
Learning Objectives
- Compare and contrast the life cycles of angiosperms (flowering plants), gymnosperms (conifers), non-seed vascular plants (ferns), and nonvascular plants (mosses)
- Describe the structures and functions of the flower, seed, and fruit in the angiosperm life cycle
- Describe the process, locations, and significance of angiosperm gametogenesis and fertilization, including double fertilization
- Predict mechanisms of pollination based on flower characteristics
- Describe the process and significance of seed maturation, dormancy, and germination
- Predict mechanisms of seed dispersal based on fruit characteristics
Sexual Reproduction in Plants: Alternation of Generations
The text below is adapted from OpenStax Biology 32.1
Plants have two distinct multicellular stages in their life cycles, a phenomenon called alternation of generations (in contrast to the haplontic and diplontic life cycles). These two stages are the multicellular haploid gametophyte and the multicellular diploid sporophyte. This life cycle is very different from most types of animal reproduction where there is only one multicellular stage: a diploid organism which produces single-celled haploid gametes.
Before we revisit this life cycle, a reminder of some terms:
- Gamete: a mature haploid male or female germ cell that is able to unite with another of the opposite sex in sexual reproduction to form a zygote
- Spore: a typically one-celled, reproductive unit capable of giving rise to a new individual without sexual fusion
Gametes are always haploid, and spores are usually haploid (and spores are always haploid in the plant alternations of generations life cycle).
In the alternation of generations life cycle, illustrated below, there is a mature multicellular haploid stage and a mature multicellular diploid stage. The multicellular haploid stage (aka the gametophyte) produces gametes via mitosis which fuse to form a diploid zygote. The zygote develops into a mature multicellular diploid individual (aka the sporophyte), which produces haploid spores via meiosis. The haploid spores then develop into a mature multicellular haploid individual.
Notice that the multicellular life stages are named for what they produce, not what they come from. The gametophyte makes gametes, and the sporophyte makes spores.
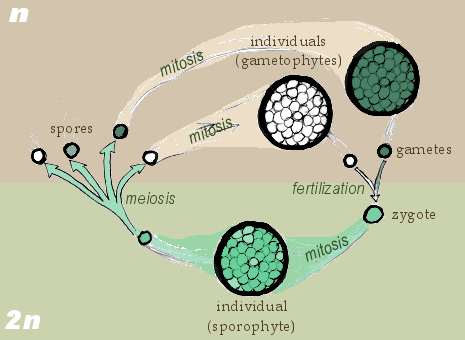
Though all plants display an alternation of generations life cycle, there are significant variations in different lineages of plants, consistent with their evolutionary history:
- In seedless non-vascular plants, or bryophytes (mosses), the haploid gametophyte is larger than the sporophyte (i.e., the plant structure that you see is the gametophyte); this is a gametophyte-dominated life cycle. The sporophyte is attached to and dependent on the gametophyte. (By “dominated” we mean “the stage of the plant you can see by eye.”)
- In seedless vascular plants (ferns), the sporophyte is larger than the gametophyte (the plant structure that you see is the sporophyte), but the gametophyte is free-living and independent from the diploid sporophyte.
- Both seedless non-vascular plants and seedless vascular plants are dependent upon water for reproduction; they produce flagellated sperm that must swim to the egg.
- The life cycles of gymnosperms (conifers) and angiosperms (flowering plants) are dominated by the sporophyte stage (the plant structure that you see is the sporophyte), with the gametophyte remaining attached to and dependent on the sporophyte (reverse of bryophytes). This is a sporophyte-dominated life cycle.
- Both gymnosperms and angiosperms produce pollen which delivers sperm to eggs without water, and seeds which protect the embryo for dispersal.
Though they both have sporophyte-dominated life cycles, gymnosperms and angiosperms differ in that:
- gymnosperms do not have flowers, do not have double fertilization, and have “naked” seeds,
- angiosperms have flowers, have double fertilization, and have fruit-covered seeds (more on each of these features this later in this reading)
The video below describes reproduction in gametophyte-dominant nonvascular plants (e.g., mosses):
The video below describes reproduction in sporophyte-dominant vascular plants (e.g., gymnosperms and angiosperms):
Reproduction in Angiosperms
The information below is adapted from OpenStax Biology 32.1
We’ll look more closely at reproduction in angiosperms, which are unique among plants for three defining features: they have flowers which are commonly used to attract animal pollinators, they reproduce via a process called double fertilization, and they have fruit-covered seeds to facilitate seed dispersal.
- Flowers are adaptations to attract pollinators
- Double fertilization is an adaptation to invest resources for nourishment of the developing embryo, in a unique way compared to other plants
- Fruits are adaptations to facilitate seed dispersal
All the information below is specific to angiosperms, unless otherwise noted.
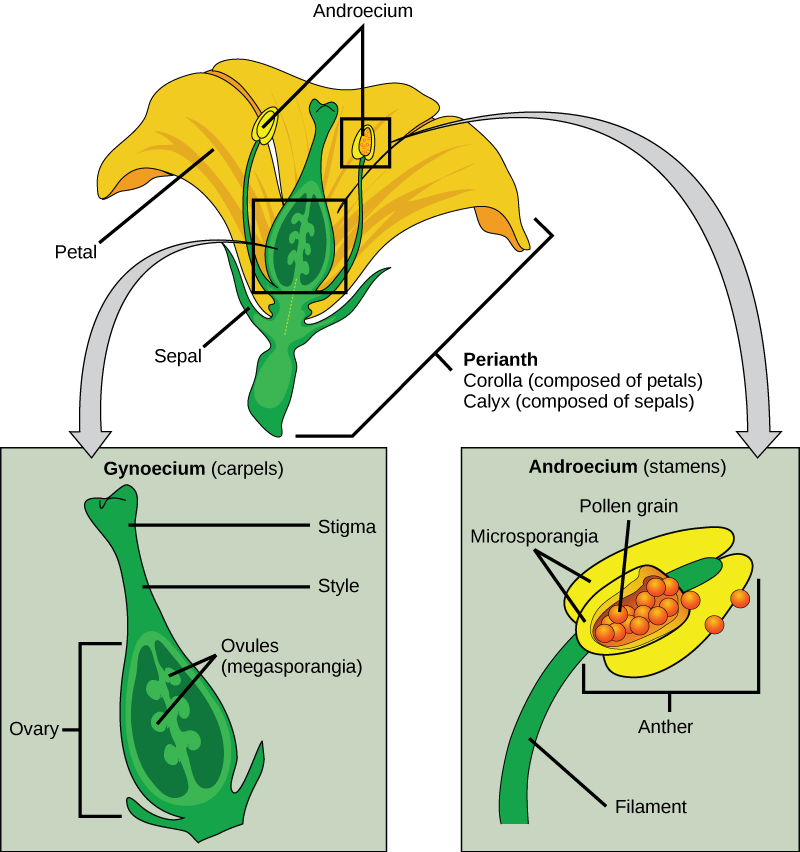
Flower Structure
A typical flower has four “layers,” illustrated and described below from external to internal structures:
- The outermost layer consists of sepals, green, leafy structures which protect the developing flower bud before it opens.
- The next layer is comprised of petals, modified leaves which are usually brightly colored, which help attract pollinators.
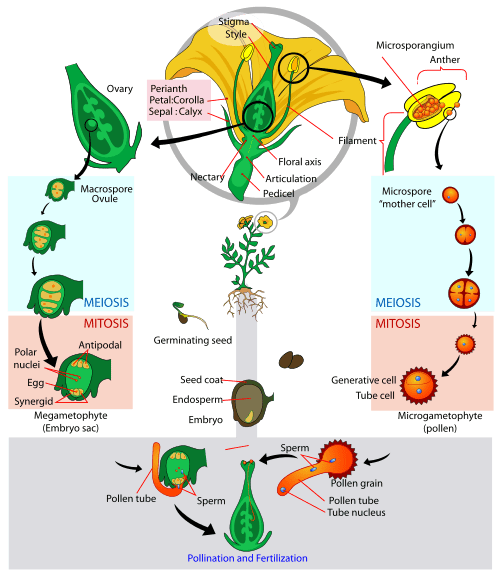
- The third layer contains the male reproductive structures: the stamens; stamens are composed of anthers and filaments. Anthers contain the microsporangia, the structures that produce the microspores, which go on to develop into male gametophytes. Filaments are structures that support the anthers.
- The innermost layer contains one or more female reproductive structures: the carpel; each carpel contains a stigma, style, and ovary. The ovaries contain the megasporangia, the structures that produce the megaspores, which go on to develop into female gametophytes. The stigma is the location where pollen (the male gametophyte) is deposited by wind or by pollinators. The style is a structure that connects the stigma to the ovary.
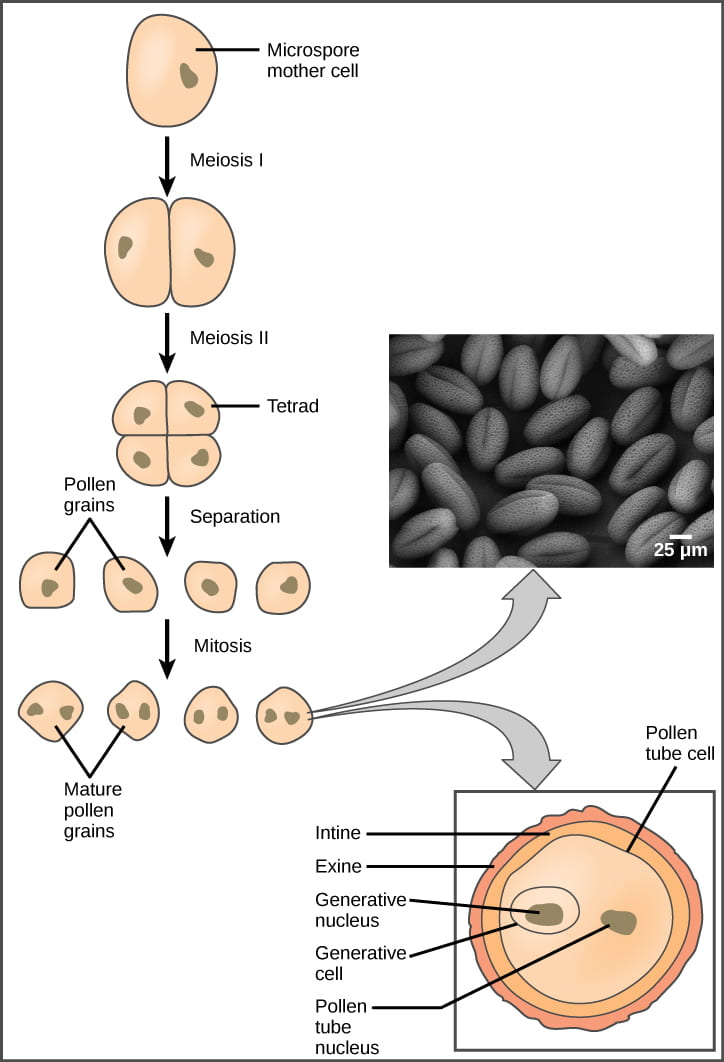
The Male Gametophyte: the Pollen Grain
Pollen is the male gametophyte in angiosperms and gymnosperms. Pollen is often described in everyday language as plant sperm, but this is not accurate! Pollen is not sperm (a gamete); pollen is the male gametophyte: a multicellular, haploid organism that produces the sperm.
Due to its protective covering that prevents desiccation (drying out) of the sperm, pollen is an important adaptation in facilitating colonization of land by plants during early plant evolution. Pollen allows angiosperms (and gymnosperms) to reproduce away from water, unlike mosses and ferns which require water for sperm to swim to the female gametophyte.
Pollen development in angiosperms occurs in a structure called the microsporangium (micro = small), located within the anthers. The microsporangia (plural of microsporangium) are pollen sacs in which the microspores develop into pollen grains.

Microspores are haploid, like all spores the alternation of generations life cycle a spore; but microspores are derived from a diploid cell. Within the microsporangium, the diploid microspore mother cell divides by meiosis to give rise to four microspores, each of which will ultimately form a pollen grain, illustrated below. This process is similar to production of gametes in animals (but notice that haploid gametes in plants are produced by mitosis from a haploid gametophyte). Upon maturity, the microsporangia burst, releasing the pollen grains from the anther where they have the opportunity to be transported to stigmas by wind, water, or an animal pollinator (more on pollinators below).
Mature pollen grains contain two cells (recall that pollen is multicellular!): a generative cell and a tube cell. The small generative cell is contained within the larger tube cell. When the pollen grain reaches the stigma of a flower, it undergoes a process called germination (not the same as seed germination). During pollen germination:
- The tube cell begins growing into the flower stigma, through the style, and down to the bottom of the ovary. As the tube cell grows, it forms a pollen tube: a conduit to deliver sperm to the egg.
- The generative cell migrates through the pollen tube to the ovary for fertilization (recall that the generative cell is inside of the tube cell). While traveling inside the pollen tube, the generative cell divides to form two sperm cells. Both sperm cells are required for successful fertilization in angiosperms in a process called double fertilization (more on that below).
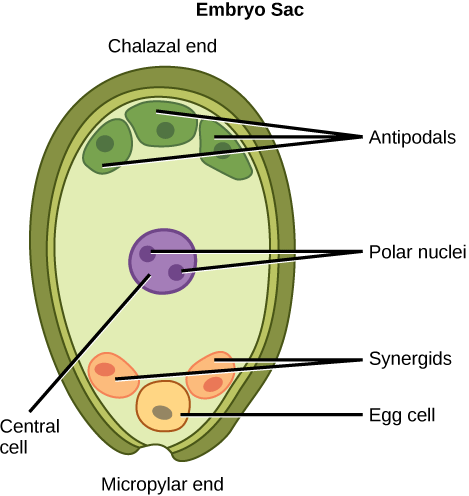
The Female Gametophyte: the Embryo Sac
The general development of the female gametophyte, or embryo sac, has two distinct phases. First, a single cell in the diploid megasporangium (mega = large), located within the ovules, undergoes meiosis to produce four megaspores. Only one megaspore survives (again similar to gamete production in animals).
In the second phase of female gametophyte development, the surviving haploid megaspore undergoes mitosis without complete cell division to produce an eight-nucleate, seven-cell female gametophyte: the embryo sac, illustrated below. Within the embryo sac:
- Two of the nuclei (the polar nuclei) move to the center of the embryo sac and fuse together, forming a single, diploid central cell. This central cell later fuses with a sperm cell to form the triploid endosperm, which will ultimately provide nourishment for the developing embryo (analogous to yolk in animal eggs).
- Three nuclei position themselves on one side of the embryo sac and develop into the antipodal cells, which later degenerate to provide nourishment to the embryo sac.
- On the opposite side of the embryo, the nucleus closest to the micropyle (the site where sperm enter the embryo sac) becomes the female gamete, or egg cell.
- The two nuclei on either side of the egg cell will develop into synergid cells. The synergids use chemical signaling to help guide the pollen tube for successful fertilization.
Once fertilization is complete, the resulting diploid zygote develops into the embryo, and the fertilized ovule forms the other tissues of the seed. The ovule wall will become part of the fruit (more on fruits below). Keep in mind that the ovule is part of the sporophyte; in contrast, the structures within the embryo sac are produced by the gametophyte.
Pollination Syndromes: how Pollen gets to the Embryo Sac
It may sound like a disease, but pollination “syndrome” just means the way a particular plant species is pollinated. The majority of pollinators are animals, including insects (like bees, flies, and butterflies), bats, or birds. Some plant species are pollinated by abiotic agents, such as wind and water. Plants that are pollinated by animals must either produce nectar to attract and feed the animals, or extra pollen that is eaten by the animals. Plants that are pollinated by wind or water must produce massive quantities of pollen since the probability of the pollen landing on a stigma of the right species is low (wind and water pollination is analogous to broadcast spawning). The mechanism of pollination and the features of the flower are tightly linked:
- Colored, highly scented flowers tend to be pollinated by bees, butterflies, wasps, or flies. These insects are active during the day and are able to detect bright colors and have a strong sense of smell. Different smells attract different pollinators, with sweet smells attracting bees and butterflies, and rotting smells attracting flies. Many insect-pollinated flowers have additional color patterns in the UV range, which insects are capable of seeing while humans cannot.
- White or pale-colored, highly scented flowers tend to be pollinated by moths and bats which are active at night. The light coloring makes them easier to see at night, and they tend to smell musky or fruity. Flowers pollinated by bats are larger than those pollinated by moths.
- Brightly colored, odorless flowers tend to be pollinated by birds, which do not have a strong sense of smell. The flowers tend to have a curved, tubular shape to accommodate the bird’s beak.
- Small green, petal-less flowers tend to be pollinated by wind. Wind-pollinated flowers do not produce nectar, but they must produce excessive quantities of pollen. Gymnosperms such as pines, which do not have flowers, are also pollinated by wind.
- Some aquatic plants are pollinated by water; the pollen floats and the water carries it to another flower.
Some examples of different pollination syndromes are shown below:

And this video briefly describes the different pollination syndromes listed above:
Double Fertilization: the Embryo and the Endosperm
The text below was adapted from Openstax Biology 32.2
The phenomenon of double fertilization, or two fertilization events, is unique to angiosperms and does not occur in any other type of plant or other organism.
As described above, after pollen is deposited on the stigma, it germinates and grows through the style to reach the ovule. The pollen tube cell grows to form the pollen tube, guided to the micropyle by chemical signals from the synergid cells. The generative cell travels through the tube to the egg and divides mitotically to form two sperm cells. One sperm fertilizes the egg cell, forming a diploid zygote; the other sperm fuses with the two polar nuclei, forming a triploid cell that develops into the endosperm, which serves as a source of nutrition for the embryo during development, and also after germination. Together, these two fertilization events in angiosperms are known as double fertilization, illustrated below. After fertilization is complete, no other sperm can enter. The fertilized ovule forms the seed, and the ovary becomes the fruit, usually surrounding the seed.

The image below puts each of these steps in context with each other:
This video gives a simplified (but very engaging) overview of double fertilization, as well a nice review of flower structure:
Seed Maturation: Development, Desiccation, and Dormancy
Just like the evolution of pollen, the evolution of the seed in gymnosperms and angiosperms was an important adaptation allowing plants to colonize land away from water due to the protection of the embryo within the plant. (Thus the seed is analogous to the amniotic egg in animal reproduction.)
After fertilization in angiosperms, the zygote enters a temporary period of development (shown below). It first divides to form two cells: the upper cell, or apical cell, and the lower cell, or basal cell.
- The division of the basal cell gives rise to the suspensor, which eventually makes connection with the maternal tissue. The suspensor does not become part of the future plant, but instead provides a route for nutrition to be transported from the mother plant to the growing embryo. (The suspensor is a type of “extra-embryonic” tissue and is analogous to the umbilical cord in placental mammals.)
- The apical cell also divides, giving rise to the embryo: the actual embryo that will develop into a plant.
- At the same time, the endosperm accumulates starches, lipids, and proteins, and then nourishes the developing cotyledons (embryonic leaves). The cotyledons will serve as a nutrient and energy store for embryo development after germination (before the plant can photosynthesize to produce its own supply of carbon).
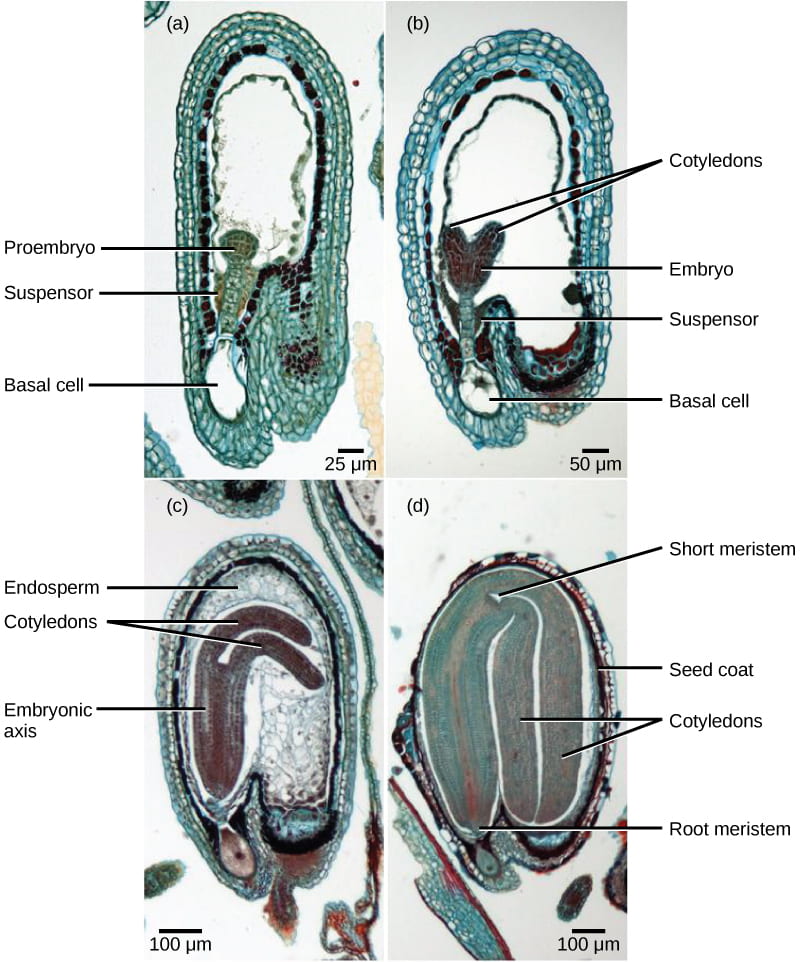
After completing this early developmental period, the seed then undergoes desiccation where it loses up to 95% of its water; this water loss which dehydrates the seed, causing extremely low metabolic activity, and “concentrating” the seed’s sugars to protect the cells from freezing during winter months (extremely high sugar concentration is a natural antifreeze).
Desiccation causes embryonic development to be paused, and the seed enters a period of dormancy. Dormancy is a period of no growth and minimal (almost nonexistent) metabolic activity which can last months, years, or even centuries in some cases.
Dormancy allows time for dispersal, and growth is resumed only once the seed detects that conditions are appropriate for seedling growth. In these conditions, the seed will then germinate or re-initiate development. Once development is reactivated, the developing seedling will rely on the food reserves stored in the cotyledons until the first set of leaves begin photosynthesis. The signal to initiate seed germination is indicator that conditions are favorable for growth and, depending on the species, can include:
- water, indicating the start of the rainy season and re-hydrating the seed
- specific wavelengths of light, indicating favorable sunlight conditions necessary for photosynthesis and the seed is not buried too far under the soil
- a sustained period of cold, indicating that the seed does not germinate until the cold season is over
- fire, typical of forest trees, indicating reduced competition from existing tall tree
- scarification, or chemical treatment with acids, indicating that the seed has passed through the digestive tract of an animal
Fruit and Seed Dispersal
After fertilization, the ovary of the flower develops into the fruit. While we tend to think of fruits as being sweet, biologically a fruit is any structure that develops from an ovary after fertilization. The biological purpose of the fruit is seed dispersal, allowing the seed to be spread far from the mother plant, so they may find favorable and less competitive conditions in which to germinate and grow.
Some fruits have built-in mechanisms so they can disperse by themselves, whereas others require the help of agents like wind, water, and animals. As with pollination syndromes and flower structure, you can often predict a fruit’s dispersal mechanism based on structure, composition, and size:
- Propulsion-dispersed fruits, such as violets, actually “explode” out of the plant.
- Wind-dispersed fruits, such as dandelions, are lightweight and may have wing or parachute-like appendages that allow them to be carried by the wind.
- Water-dispersed fruits, such as coconuts, are light or buoyant, giving them the ability to float.
- Animal-dispersed fruits may be either have tiny “hooks” all over them so that they attach to passing animals and later fall off in a new location (like sandburs), or very sweet or fatty so that they will be eaten and deposited in a new location in the feces (like blackberries). Fruits dispersed by birds tend to be brightly colored as birds have a highly developed sense of sight; fruits dispersed by mammals tend to be highly scented as mammals have a highly developed sense of smell.

