
This page is under revision
Learning Objectives
- Place and identify the clade Chordata on a phylogenetic tree within the domain Eukarya and the kingdom Animalia
- Identify and describe the key adaptations of all chordates
- Identify and describe key adaptations present in vertebrate lineages, and use these adaptations to differentiate among major vertebrate groups
- Explain how specific adaptations allowed different vertebrate taxa to exploit increasingly dry environments
- Place the evolution of the major vertebrate taxa in chronological order and identify key geologic time points in their evolution
Chordates on the Phylogenetic Tree of Life
Chordates, or animals with a notochord, form a clade within the Deuterostomes. Deuterostomes are triploblastic animals where the anus (as opposed to the mouth) is the first part of the digestive tract to develop during embryogenesis. Deuterostomes include two only two phyla: Chordates and Echinoderms, including starfish, sea urchins, sand dollars, etc. (You can review information on Deuterostomes in the discussion of Invertebrates.)
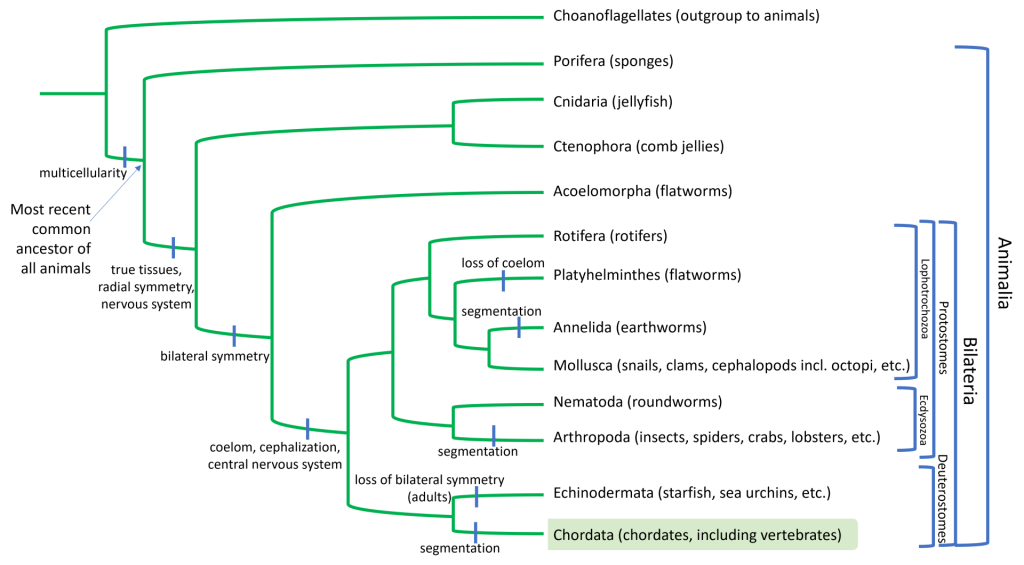
Evolutionary Adaptations Shared by all Chordates
The information below is adapted from OpenStax Biology 2e 29.1, OpenStax Biology 29.3, OpenStax Biology 29.4, , OpenStax Biology 29.6, CK-12 Biology Advanced Concepts 16.7 Vertebrate Evolution – Advanced, CK-12 Biology for High School 12.5 Vertebrate Evolution

Chordates include both invertebrate and vertebrate species, but all vertebrates share the following traits at some point during their developmental/larval or adult stages:
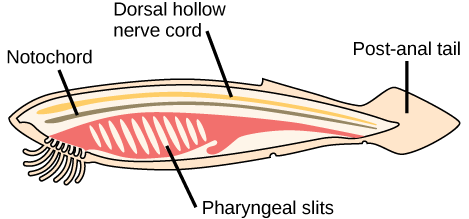
- Notochord: Chordates are named for the notochord, a flexible, rod-shaped structure that arises from mesodermal tissue during embryonic development. The notochord is found in the embryonic stage of all chordates and in the adult stage of some chordate species. It is located between the digestive tract and the nerve cord, and it provides support throughout the length of the body.
- Dorsal hollow nerve cord: The dorsal hollow nerve cord arises from ectodermal tissue that rolls into a hollow tube during development. In chordates, it is located dorsal (along the back) from the notochord. In contrast, other animal phyla have solid (rather than hollow) nerve cords that are located either ventrally (along the belly) or laterally (on the sides). The nerve cord found in most chordate embryos develops into the brain and spinal cord, which compose the central nervous system.
- Pharyngeal slits: The pharyngeal slits are openings in the pharynx (the region just behind the mouth) that extend to the outside environment. In organisms that live in aquatic environments, pharyngeal slits allow for the exit of water that enters the mouth during feeding.
- Some invertebrate chordates use the pharyngeal slits to filter food out of the water that enters the mouth.
- In vertebrate fishes, the pharyngeal slits are modified into gill supports and/or jaw supports.
- In tetrapods (amphibians, reptiles, birds, and mammals), the slits are modified into components of the ear and tonsils.
- Muscular post-anal tail: The muscular post-anal tail is a posterior elongation of the body, extending beyond the anus. The tail contains skeletal elements and muscles which provide a source of locomotion in aquatic species. In some terrestrial vertebrates, the tail also helps with balance, courting, and signaling when danger is near. In apes and humans, the post-anal tail is vestigial, meaning it is reduced in size and nonfunctional.
The image below shows these traits on a cartoon of a generalized chordate:
Evolutionary Adaptations among Chordate and Vertebrate Lineages
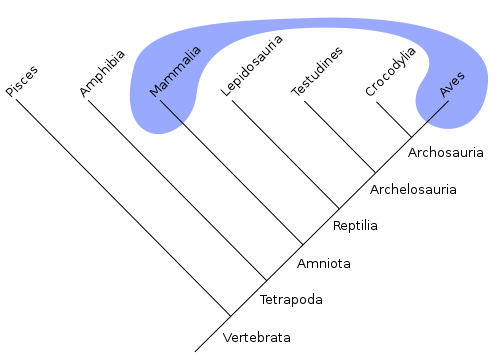
Chordates make up just over 3% of all animal species, but they include the largest animal species on Earth. Below is a phylogenetic tree visualizing chordate evolutionary relationships. Viewing the tree, note that not all chordates are vertebrates! There are three groups of non-vertebrate chordates: subphylum Cephalochordata (lancelets), subphylum Urochordates (tunicates), and class Myxini (hagfish).

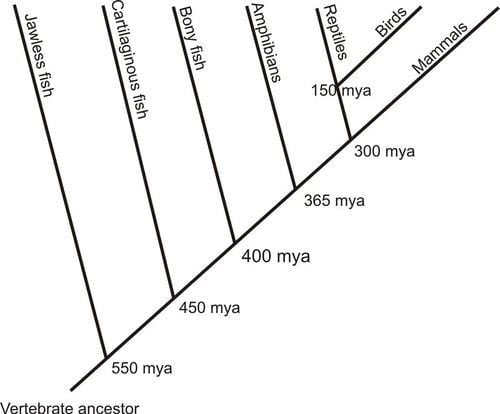
All chordates possess the four traits described above: notochord, dorsal hollow nerve chord, pharyngeal slits, and a muscular post-anal tail. In addition, they are triploblastic deuterostomes that evolved from an ancestor with cephalization (see the reading on Invertebrate Animals for a review of these concepts). New adaptations arose in different chordate lineages as they diverged over time, including a cranium (skull), vertebral column, jaw, bony skeleton, lungs, lobed fins, four limbs, amniotic egg, endothermy, feathers, wings, hair, and milk production. In the section below, we will discuss the evolution of different chordate groups with emphasis on these evolutionary adaptations.
Before we walk through these details, here is the Crash Course take on Chordates (please note, as entertaining as Hank Green may be, there is no such thing as a “living fossil”):
- The cranium (skull) evolved in a group of chordates called craniates (Craniata):
- Fossils of the earliest known craniate date to approximately 518 MYA during the Cambrian explosion; these animals were approximately 1 inch long, and had a defined skull with a distinct head and tail, but no evidence of a vertebral column.
- Modern day hagfish (Myxini) are non-vertebrate chordates that are the only living species with a skull but no vertebral column. Hagfish do have some vertebrae-like structures and there is some evidence that they may have evolved from a vertebra-bearing ancestor, but lost this trait. The hagfish skeleton consists of the skull and the notochord. Hagfish are jawless.

- The vertebral column evolved in an early craniate, resulting in the first vertebrate. The presence of the vertebral column is the defining evolutionary feature of this group. In adult vertebrates, the vertebral column replaces the notochord, which is only seen in the embryonic stage. Fishes were the earliest vertebrates, with jawless species being the earliest and jawed species evolving later. Jawless fishes are called agnathostomes (“a” = “without” and “gnathos” = “jaw”)
- One of the earliest known possibly-vertebrate species is represented by a single known fossil approximately 1 in long and dating to approximately 518-490 MYA, although it is not universally accepted that this fossil represents a true vertebrate.
- Living jawless fishes (agnathostomes) such as lampreys (Hyperaortia), are craniates with a vertebral column that represent a vertebrate linage that diverged from other vertebrate lineages over 500 million years ago. Lampreys lack fins, causing them to look tubular.
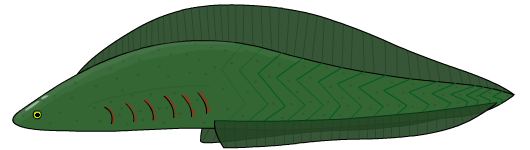
- Evolution of the jaw occurred in a group of vertebrates called gnathostomes or ‘jaw-mouths‘; a jaw is defined as a hinged structure attached to the cranium that allows an animal to grasp and tear its food. Evolution of the jaw approximately 430 MYA was one of the most significant developments in early vertebrate evolution, facilitating an evolutionary arms race between predators, competitors, and prey. In addition to jaws, early gnathostomes also possessed two sets of paired fins, allowing the fishes to maneuver accurately and become mobile predators. These two traits allowed early gnathostomes to exploit food resources that were unavailable to jawless fishes. Most modern fishes are gnathostomes (jawed).
- A famous example of early gnathostomes are the placoderms, which were a diverse group of jawed fish with an abundance of fossils dating from approximately 430 – 360 MYA. Placoderms were large (up to ~29 feet long) predators with bony plates forming armor over their heads, but their internal skeletons were typically composed of cartilage (with one noteworthy exception discovered in 2020).
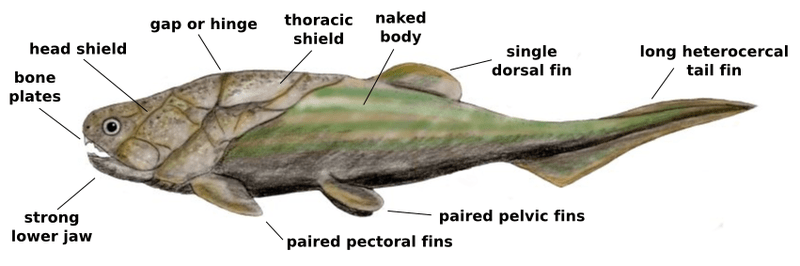
- Cartilaginous skeletons appear in the cartilaginous fish (Chondrichthyes), living jawed fishes (gnathostomes) that possess paired fins and a skeleton made of cartilage, including sharks, rays, skates, sawfishes and a few dozen species of fishes called chimaeras (ghost sharks). Cartilaginous fish first appear in the fossil record approximately 395 MYA and may have evolved from one group of early placoderms. Some recent fossil evidence suggests that the possibly-placoderm ancestors of cartilaginous fish once had bony endoskeletons but lost this trait over evolutionary time, replacing their bony skeleton with a skeleton of cartilage; this hypothesis is currently under debate in the scientific community.
- Mineralized (bony) skeletons evolved in bony fishes (Osteichthyes), appearing in the fossil record approximately 420 MYA. Bony fish may have evolved from another group of placoderms, distinct from the ancestors of cartilaginous fish. The vast majority of living fishes belong to this group, which consists of approximately 30,000 species, making it the largest class of vertebrates in existence today. The ‘bony fish’ group includes the ray-finned fishes Actinopterygii), and lobe-finned fishes (Sarcopterygii).
- Lungs first appeared in early bony fish; these lungs later evolved into swim bladders which are present in modern-day fish (NOT the other way around). What were lungs doing in ancient bony fish? There are a number of hypotheses, but one of the most prominent is that early lungs allowed fish to “gulp” air in oxygen-poor conditions; this air could then supplement oxygen obtained from gills and circulated in the circulatory system.
- Lobed fins were an were an important adaptation that appeared in the lobe-finned fishes (Sarcopterygii) approximately 425 MYA; lobed fins allowed for later evolution of tetrapods (four-legged vertebrates). Lobed fins differ from the fins of ray-finned fishes in that the fin extends from a fleshy, lobe-like structure that resembles a limb bud and is connected to the body by a single bone. Most lobe-finned fishes are now extinct, but living species include coelacanths and lungfishes.
Epipelagic
- Four limbs first appeared in a group of early tetrapods (four-limbed vertebrates) nearly 400 million years ago in a now-extinct amphibian species. Evolution of tetrapods from lobe-finned freshwater fishes represented a significant change in body plan from one suited to organisms that respired and swam in water, to organisms that breathed air and moved onto land; these changes occurred over a span of 50 million years.
- Tiktaalik is a famous fossil of a “tetrapod-like fish” that lived in a shallow water environment about 375 million years ago. Tiktaalik seems likely represent an intermediate form between fishes having lobe fins and tetrapods having limbs.
- The earliest known true tetrapods include Ichthyostega and Acanthostega. Both were aquatic species (not truly terrestrial), and they may have had both gills and lungs. Though they had true limbs, their limbs could not be pulled in under the body and would not have supported their bodies well out of water. They probably lived in shallow freshwater environments, and may have taken brief terrestrial excursions.
Watch the video below about the importance of the fossil Tiktaalik to our understanding of tetrapod evolution:
- The first tetrapods were amphibians (Amphibia). Modern-day amphibians include frogs, salamanders, and caecilians. The term amphibian loosely translates from the Greek as ‘dual life’ which is a reference to the metamorphosis that many frogs and salamanders undergo and their mixture of aquatic and terrestrial environments in their life cycle. Most amphibian species must remain forever associated with a moist environment, at a minimum in early development as eggs. In addition to breathing with lungs, amphibians also exchange gas through their moist and permeable skin, which is kept moist through mucus glands in the skin.
- Early amphibians were predators who dominated the terrestrial landscape, but were limited to wet and swampy areas. Early amphibians grew as large as 18 feet in length.

- The amniotes (Amniota) evolved approximately 300 to 360 MYA from a group of amphibians with the origin of the amniotic egg. Modern-day amniotes include reptiles, birds, and mammals. The amniotic egg is protected by amniotic membranes (fluid-filled membranes which function in embryonic development), with a solid eggshell surrounding and protecting the membrane-enclosed embryos. The evolution of the amniotic egg approximately 300 to 360 million years ago was key to allowing tetrapods to reproduce on land and marked the beginning of a flourishing population of terrestrial tetrapods.
- There are many lineages of amniotes (including many extinct groups), but here we will emphasize two important groups of amniotes: the sauropsids (ancestors of lizards, snakes, turtles, crocodilians, dinosaurs, and birds), and the synapsids (ancestors of mammals).
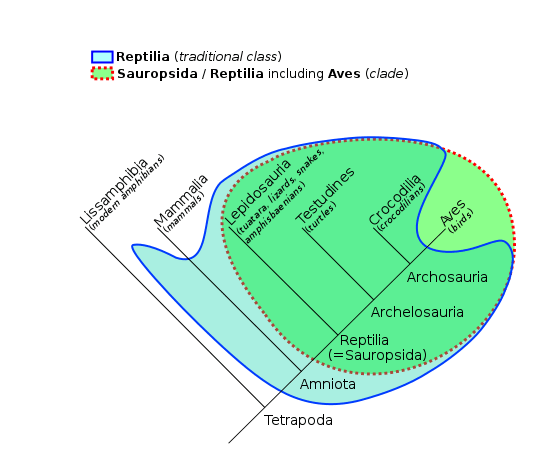
Maksim Dolgun. Based on File:Phylogenetic-Groups.svg by TotoBaggins at English Wikipedia.
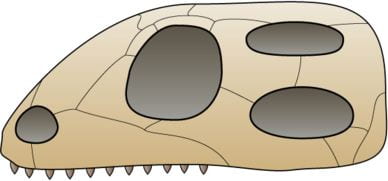
- The earliest reptiles (Reptilia, within the Sauropsidia) appear in the fossil record approximately 320 MYA, and the earliest known true reptile was a small lizard-like animal approximately 8-10 inches long (including its tail). Unlike modern reptiles, this early reptile was an anapsid, meaning it had a solid skull without any holes (or “temporal openings”) on the side. This skull structure is important because it affects the way muscles connect from the jaw to the skull, influencing jaw strength and movements; having no holes means smaller jaw muscles and thus less jaw strength.

- Reptiles were the dominant terrestrial vertebrates from approximately 250-66 MYA, and a clade of sauropsids called diapsids are the ancestors of lizards, snakes, turtles, crocodilians, dinosaurs, and birds. Diapsids are defined by the presences of two temporal openings (holes) in each side of the skull; this structure allows for attachment of larger, stronger jaw muscles and wider jaw opening, compared to amniotes lacking these holes in the skull.
- Reptiles use lungs to breathe and have scaly skin containing the protein keratin and waxy lipids, which reduced water loss from the skin. This thick, impermeable skin means that reptiles cannot use their skin for respiration, like amphibians, and thus all breathe with lungs.
- Reptiles, like fish and amphibians, are ectothermic, meaning their main source of body heat comes from the environment. In addition to being ectothermic, reptiles are categorized as poikilotherms, or animals whose body temperatures vary rather than remain stable. Reptiles have behavioral adaptations to help regulate body temperature, such as basking in sunny places to warm up and finding shady spots or going underground to cool down. The advantage of ectothermy is the low metabolic energy needed to survive (e.g., about 10% of a similarly-sized endotherm).
- Birds (Aves, within Sauropsidia) are the only group of dinosaurs that are still living. Birds evolved approximately 100 to 150 MYA from a group of dinosaurs called theropods. (Even though birds descended from dinosaurs, which are reptiles, we will consider birds as a group distinct from reptiles for the purpose of this discussion with the understanding that this designation does not accurately reflect phylogenetic history and relationships.)
- One important fossil of an animal intermediate between theropod dinosaurs and birds is Archaeopteryx, which is from the Jurassic period. Archaeopteryx appears to have characteristics of both dinosaurs and birds: the fossilized skeleton of Archaeopteryx looks like that of a dinosaur, and it had teeth whereas birds do not, but it also had feathers modified for flight, a trait associated only with birds among modern animals. Fossils of older feathered dinosaurs exist, but the feathers do not have the characteristics of flight feathers.
- The most obvious characteristic that sets birds apart from other modern vertebrates is the presence of feathers, which are modified scales. In addition to allowing for flights, feathers are also very effective insulation, trapping air in spaces between each feather to decrease the rate of heat loss. Feather coloration can play roles in camouflage or social signaling.
- Birds are endothermic, meaning they produce their own body heat via their metabolism. Compared to ectothermy, endothermy requires more energy and thus more food. Because birds fly, they require large amounts of energy, necessitating an especially high metabolic rate.

- Mammals (Mammalia, within Synapsidia) evolved approximately 200-230 MYA from a group of reptile-like amniotes called synapsids. Synapsids are defined by the presence of a single hole in in each side of the skull. Similar to diapsids (ancestors of modern-day reptiles), the structure of the synapsid skull allows for attachment of larger, stronger jaw muscles and wider jaw opening, compared to amniotes lacking this hole in the skull. Mammals evolved from a group of synapsids called therapsids, and the cynodonts are the group of therapasids that include the most recent common ancestor of all mammals.
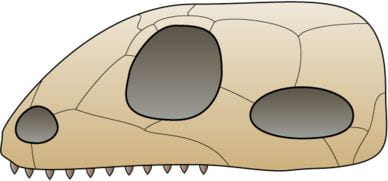
- Mammals are amniotes that retain features of the amniotic egg, even though only one group (the monotremes) still lay eggs. All mammals produce milk from mammary glands for their offspring.
- The skeletal system of mammals possesses many unique features. Unlike other vertebrates, the lower jaw of mammals consists of just one bone; the additional jaw bones found in other vertebrates have been modified to function in hearing and form the three small bones in the mammalian ear, and is one way of distinguishing fossil mammals from fossils of other synapsids. The musculature of the jaw is also unique in mammals, allowing side-to-side movement of the jaw which makes chewing possible; chewing is unique to mammals, and most mammals have different types and shapes of teeth rather than just one type and shape of tooth as in other amniotes.
- The presence of hair is one of the most obvious visual features of a mammal (even whales and dolphins have hair). Like feathers in birds, hairs are heavily modified scales; hair, feathers, and scales are homologous structures of amniotes. Like feathers in birds, hair is effective insulation, trapping air in spaces between each hair to decrease the rate of heat loss. Hair coloration can also play roles in camouflage or social signaling.
- Also like birds, mammals are endothermic, meaning they generate their own body heat via metabolic processes, and hair provides insulation to retain heat generated by metabolic work. Because the common ancestor of mammals and birds was ectothermic, endothermy evolved independently in mammals and birds; it is not a homologous trait.
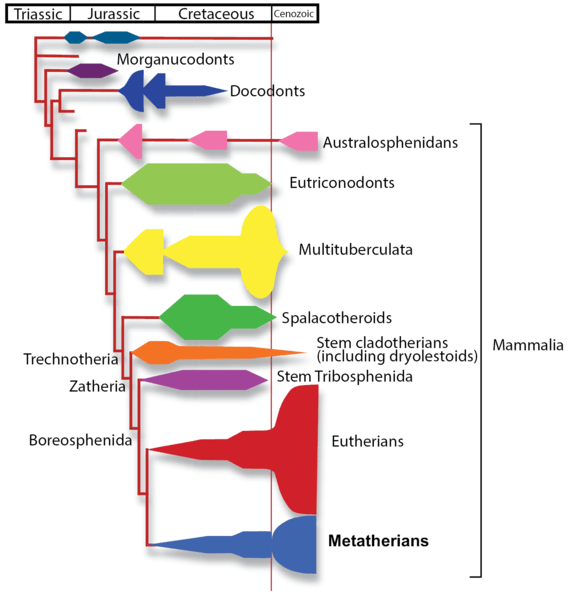
- Living mammals belong to just three clades: eutherians (including placental mammals, like us), marsupials (metatherians) which have live birth but no placenta, and the young develop within a pouch (e.g., kangaroo, opossum), and monotremes (australosphenidans) which lay eggs (platypus and echidna). Many other clades of mammals are now extinct.
Vertebrate Adaptations to Increasingly Dry Environments over Geologic Time
The story of vertebrate evolution is the story of increasing adaptation to dry environments:
- The first vertebrates were fish-like aquatic animals that lived during the early Paleozoic Era.
- The capable of leaving the water were amphibians, which evolved in the mid-Paleozoic. The mid-Paleozoic featured swampy, wet conditions that allowed amphibians to flourish. Amphibians must remain near a moist environment for their reproduction, as their eggs must be laid in water. In addition, amphibians also exchange gas through their moist and permeable skin, which is kept moist through mucus glands in the skin. The Carboniferous period in particular saw amphibian tetrapods become the dominant predators on land.
- The first amniotes evolved during the mid-Paleozoic, but were largely outcompeted by the numerous amphibians which were already occupying many of the available niches on land. As conditions became increasingly dry during the late Paleozoic, amniotes flourished due to several adaptations to dry conditions:
- the amniotic egg, which contains fluid-filled amniotic membranes surrounding the embryo and a solid shell protecting those membranes, allowed amniotes to reproduce away from water
- the more efficient lungs of amniotes meant that they did not have to rely on gas exchange through the skin to supplement gas exchange in the lungs like amphibians, which allowed for evolution of desiccation-resistant skin coverings such as scales, feathers, and hair (described below)
- the evolution of scales (in reptiles, which first appeared in the mid-Paleozoic) and then later hair (in mammals, which first appeared in the early Mesozoic) and feathers (in birds, which first appeared the late-Mesozoic) provided protection against desiccation, or drying out; in contrast, the thin, moist skin of amphibians can require access to sufficient water to prevent dehydration

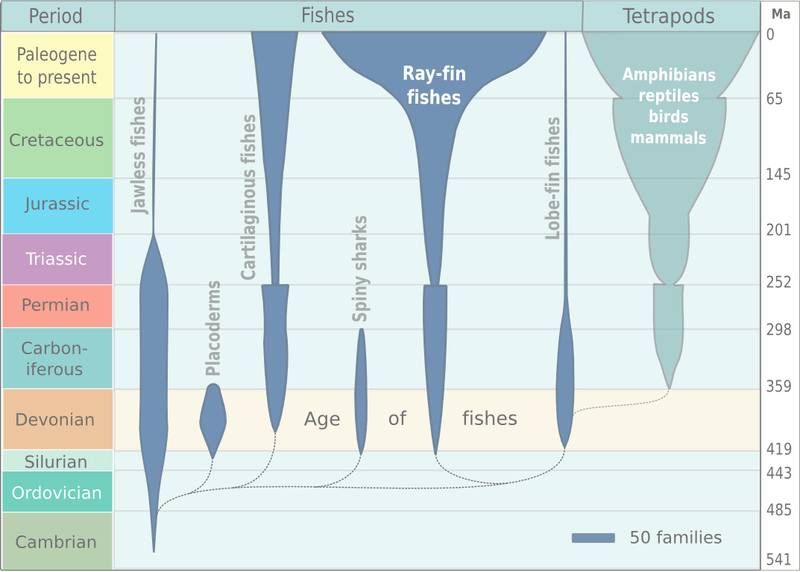
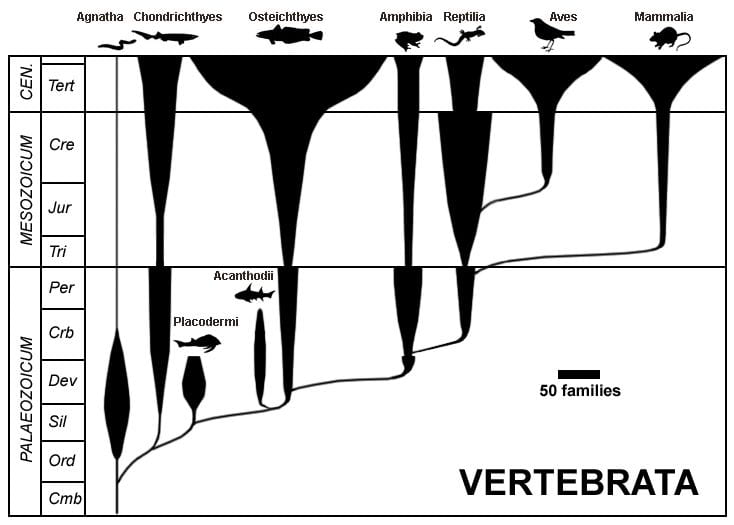
The spindle diagram below provides a visual for some of the events described above, with geologic periods of and eras of the Phanerozoic on the left.

{kind=link}
{kind=link}