
Learning Objectives
- List the sequence of early events in animal development (fertilization, cleavage, gastrulation, and organogenesis)
- Identify key structures in/on eggs and sperm, and explain their roles in fertilization
- Explain and describe the significance, steps, and features of fertilization and cleavage in early animal development
- Differentiate between intrinsic and extrinsic factors determining cell specialization
- Explain and differentiate between the roles of cytoplasmic determinants and induction in determining body axes during early animal development
The Purpose and Stages of Development
The information below was adapted from Khan Academy “Introduction to development.” All Khan Academy content is available for free at www.khanacademy.org
Your body consists of over 30 trillion cells, but you began as a single cell: a fertilized egg, or zygote. How did you become the large, organized multicellular individual that you are today? Development!
As an animal embryo develops, its cells divide, grow, and migrate in specific patterns to make a more and more elaborate body (plant cells perform differential expansion instead of migration). To function correctly, that body needs well-defined axes (such as head vs. tail aka anterior vs. posterior). It also needs a specific collection of many-celled organs and other structures, positioned in the right spots along the axes and connected up with one another in the right ways. How are all of these complex processes accomplished and coordinated? They occur via four essential stages in early animal development:
- Fertilization: the process of a single sperm cell combining with a single egg cell to form a zygote.
- Cleavage: rapid, multiple rounds of mitotic cell division where the overall size of the embryo does not increase. The developing embryo is called a blastula following completion of cleavage.
- Gastrulation: the dramatic rearrangement (movement) of cells in the blastula to create the embryonic tissue layers. These tissue layers will go on to produce the tissues and organs of the adult animal.
- Organogenesis: the process of organ and tissue formation via cell division and differentiation.
The last two stages, gastrulation and organogenesis, together contribute to morphogenesis: the biological processes that result in an organism’s shape and body organization.
For this reading, we will discuss the first two steps above, fertilization and cleavage. We will pick up with gastrulation and organogenesis in the next reading.
Development Step 1: Fertilization
The information below was adapted from OpenStax Biology 43.6
Fertilization is the process in which a single haploid sperm fuses with a single haploid egg to form a zygote. The sperm and egg cells each possess specific features that make this process possible.
The egg is the largest cell produced in most animal species; a human egg cell is approximately 16 times larger than a human sperm cell. The eggs of different species have similar features in common, including:
- Yolk: Eggs of different species contain varying amounts of yolk, the nutrients to support growth of the developing embryo.
- Jelly layer or zona pellucida: Each egg is surrounded by a jelly layer, composed of glycoproteins (proteins that have sugars stuck to them), that releases species-specific chemoattractants (chemical-attractors) that guide sperm to the egg. In mammals, this layer is called the zona pellucida. In placental mammals, a layer of follicular cells surrounds the zona pellucida.
- Vitelline envelope: The zona pellucida/jelly layer is separated from the egg by a membrane called the vitelline envelope, which is a second membrane outside of the cell’s plasma membrane.
- Cortical granules: Just underneath the egg’s plasma membrane are cortical granules, vesicles containing enzymes that will degrade the proteins that hold the vitelline envelope around the plasma membrane when fertilization occurs (more on this below).
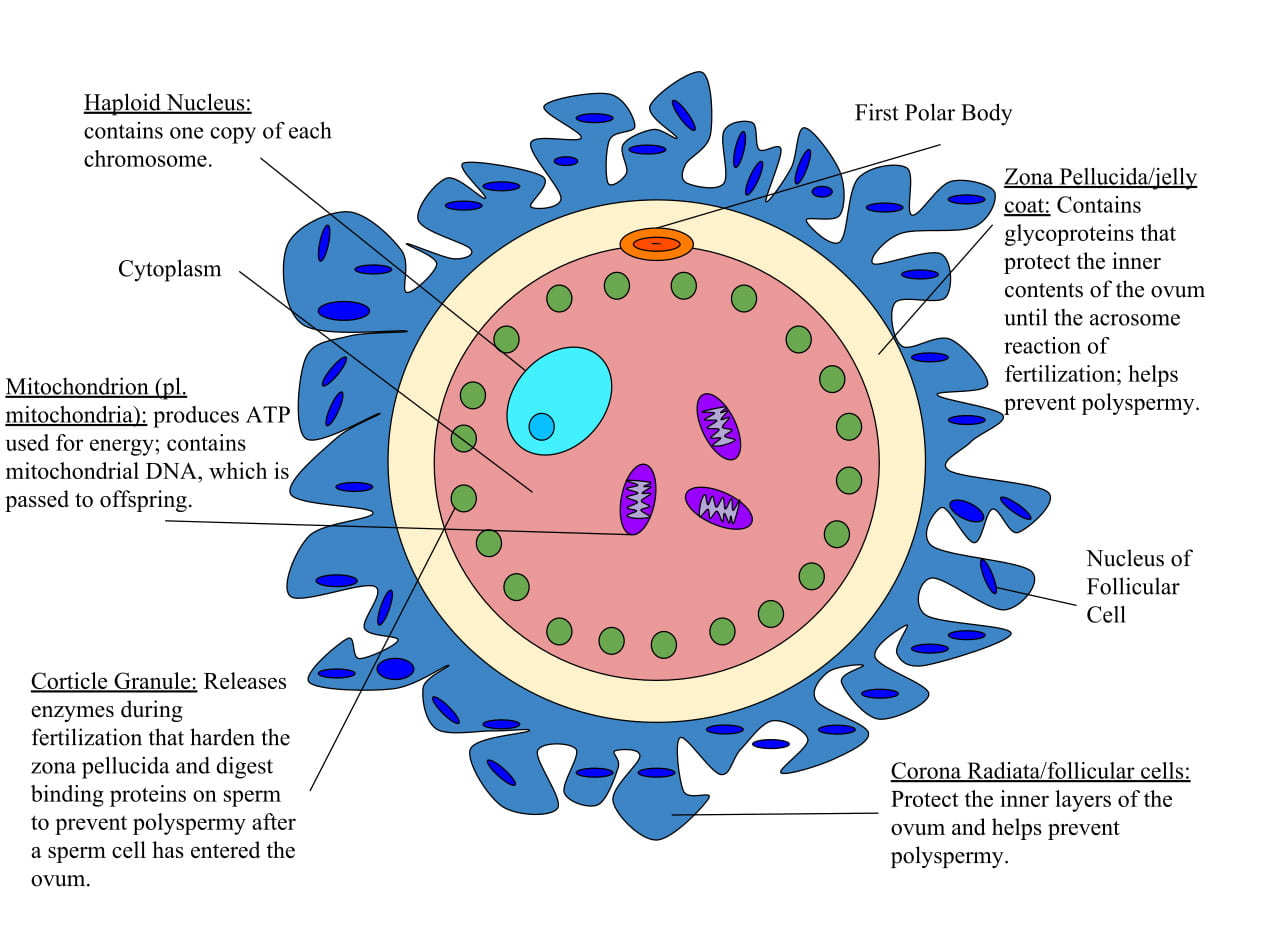
The sperm is one of the smallest cells produced in most animal species. The sperm of different species have similar features in common, including:
- Sperm structure: The sperm consists of head containing tightly packed DNA, a flagellar tail for swimming, and many mitochondria to provide power for sperm movement.
- Bindin proteins: The plasma membrane of the sperm contains proteins called bindin, which are species-specific proteins that recognize and bind to receptors on the egg plasma membrane.
- Acrosome: In addition to the nucleus, the sperm head also contains an organelle called the acrosome, which contains digestive enzymes that will degrade the jelly layer/zona pellucida to allow the sperm to reach the egg plasma membrane.

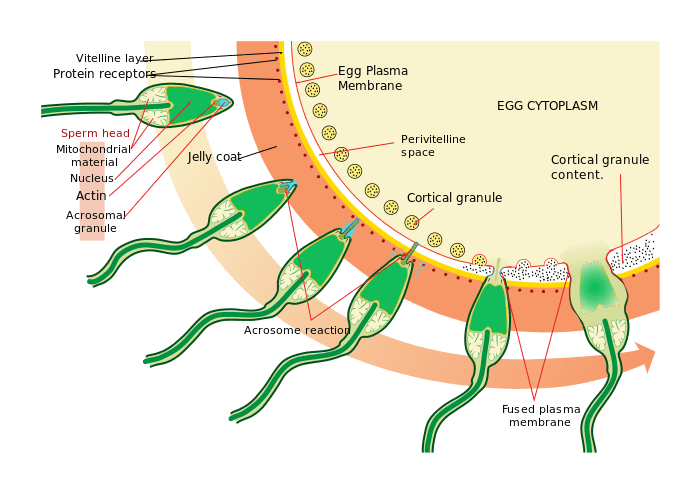
To ensure that the offspring has only one complete diploid set of chromosomes, only one sperm can fuse with each egg. Fusion of more than one sperm with an egg, or polyspermy, is genetically incompatible with life and results in zygote death. There are two mechanisms that prevent polyspermy: the “fast block” to polyspermy and the “slow block” polyspermy. These and the other steps of fertilization are described here:
- Sperm is attracted to and makes contact with the jelly layer/zona pellucida.
- The interactions between receptors on the sperm cell and glycoproteins on the egg cell initiate the acrosome reaction. Digestive enzymes are released from the acrosome in the sperm, and the enzymes destroy the jelly layer/zona pellucida to create a pathway for the sperm to reach the egg.
- The sperm reaches the egg plasma membrane, and the bindin proteins on the sperm plasma membrane contact the bindin receptors on the egg plasma membrane; this process allows the sperm and egg membranes to fuse. Bindin proteins and bindin receptors are species-specific, meaning that the sperm from one species is unlikely to be able to fertilize the egg of a different species.
- Fusion of the sperm and egg membranes initiates electrical depolarization of the entire egg plasma membrane for 10-20 seconds, temporarily preventing any other sperm from fusing with the egg plasma membrane. This membrane depolarization, mediated by an influx of sodium ions, is the fast block to polyspermy.
- The membrane depolarization then initiates a wave of calcium released across the plasma membrane.
- The calcium wave initiates the cortical reaction in the egg, where the cortical granules fuse with the egg plasma membrane, releasing digestive enzymes that degrade the bindin receptor proteins on the egg membrane. The bindin receptors have two jobs: they are sperm docking sites, and they also hold the vitelline layer against the plasma membrane.
- Destruction of the bindin receptors in the egg plasma membrane causes the vitelline layer to lift away from the egg plasma membrane, creating the fertilization envelope. The fertilization envelope is a barrier that prevents additional sperm from reaching the egg, and is the slow block to polyspermy.
- These events culminate in egg activation, causing the egg to recognize that fertilization has occurred and resulting in initiation of development.
The video below provides a nice overview of sex cells and fertilization, including the acrosome and cortical reactions:
And this video shows what happens in sea urchin eggs immediately following fertilization, including the calcium wave that initiates the cortical reaction:
Development Step 2: Cleavage and Blastula Stage
After fertilization activates the egg, the egg begins a series of rapid cell divisions called cleavage, illustrated below. Typical, non-cleavage cell division occurs every 18-24 hours, but cleavage cell divisions can occur as frequently as every 10 minutes. During cleavage, the cells divide without growing between divisions, so the large single-celled zygote divides into smaller and smaller cells called blastomeres. After the cleavage has produced over 100 blastomeres, the embryo is called a blastula. The blastula is usually a spherical layer of blastomeres that are considered to be the first embryonic tissue, the blastoderm. The blastoderm surrounds a fluid-filled or yolk-filled cavity, called the blastocoel (recall that a coelom is a body cavity). The blastocoel is absolutely essential for the next step of development, gastrulation, which we will discuss in the next reading.
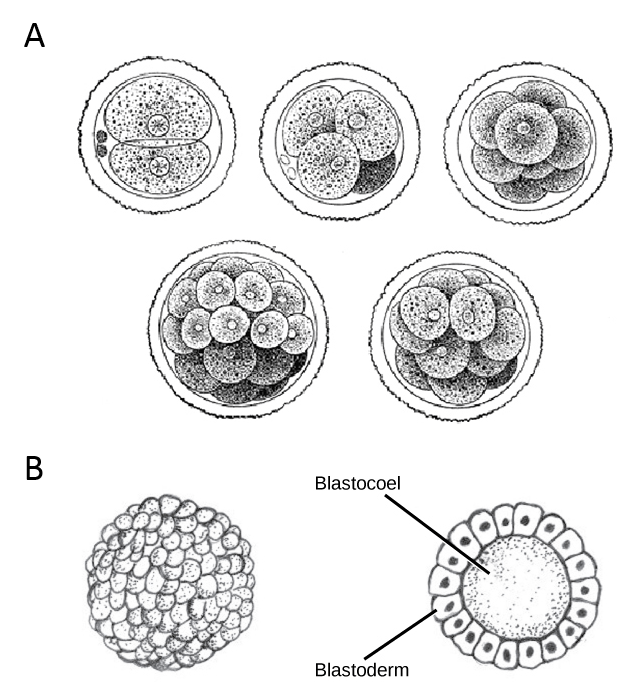
The stages of development that we’ve discussed so far are very similar across most animal lineages. But later stages of cleavage are a little different in mammals: the mammalian blastula is called a blastocyst, and, unlike the blastulas of other animal lineages, the blastocyst has an “inner cell mass” and an outer cell layer called the trophoblast. The inner cell mass will go on to form the embryo, and the trophoblast will go on to form embryonic portion of placenta. Cleavage in a placental mammal is illustrated in the diagram below.

You can visit the Virtual Human Embryo project at the Endowment for Human Development site to step through an interactive that shows the stages of embryo development, including micrographs and rotating 3-D images.
Intrinsic vs Extrinsic Factors in Cell Specialization
The information below was adapted from Khan Academy “Introduction to development.” All Khan Academy content is available for free at www.khanacademy.org
We’ve discussed 2 of the 4 general steps in early animal development. Before we move on to step number 3 (gastrulation), let’s take a moment to discuss how cells get instructions what they’re supposed to do, or what their “identify” is, in an embryo: whether it’s to divide, to migrate to a new location, or to begin specializing into a specific cell type. Cells generally get instructions from either intrinsic (internal, or “lineage”) or extrinsic (external, or “positional”) information:
- Intrinsic (internal, or lineage) information is inherited from the mother cell, as a result of cell division. For instance, a cell might inherit molecules in its cytoplasm that “tell” it that it belongs to the neural, or nerve cell-producing, lineage of the body. It inherits them because they were present in the cytoplasm of the mother cell, and thus they are present in the cytoplasm of the daughter cells. Cytoplasmic determinants, discussed in the next section of this reading, are examples of intrinsic information present in the unfertilized eggs of protostomes, before development (or fertilization) even begins.
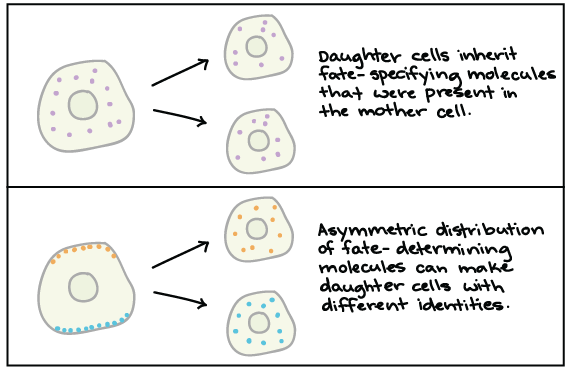
- Extrinsic (external, or positional) information is received from the cell’s surrounding environment or from neighboring cells. For instance, a cell might get chemical signals from a neighbor, instructing it to become a particular kind of photoreceptor (light-detecting neuron). Induction, discussed in the next section of this reading, is an example of an extrinsic information process that occurs in embryonic development of all animals.

Why do intrinsic and extrinsic signals matter during development? These signals are essential for the events which occur after the blastula stage of embryonic development, and (in some cases), even before that point. We’ll explore how intrinsic and extrinsic information plays a role in development in the rest of this reading and the next reading.
Determining the Body Axes: Cytoplasmic Determinants (Intrinsic Information) and Induction (Extrinsic Information)
One of the fundamental principles of animal development in all animals (except sponges) is the establishment of the body axes: animal bodies have lateral-medial (left-right), dorsal-ventral (back-belly), and anterior-posterior (head-feet) axes, illustrated below.
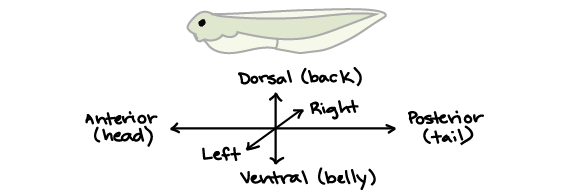
How are these axes established from a ball of apparently identical cells (the blastula)? The process is different among different lineages of animals, with body axes being heavily influenced by cytoplasmic determinants in protostomes (most invertebrates), by yolk polarity in vertebrates with large amounts of asymmetrically-distributed yolk (many fish, amphibians, reptiles, and birds), and by induction (cell-cell communication) in many mammals:
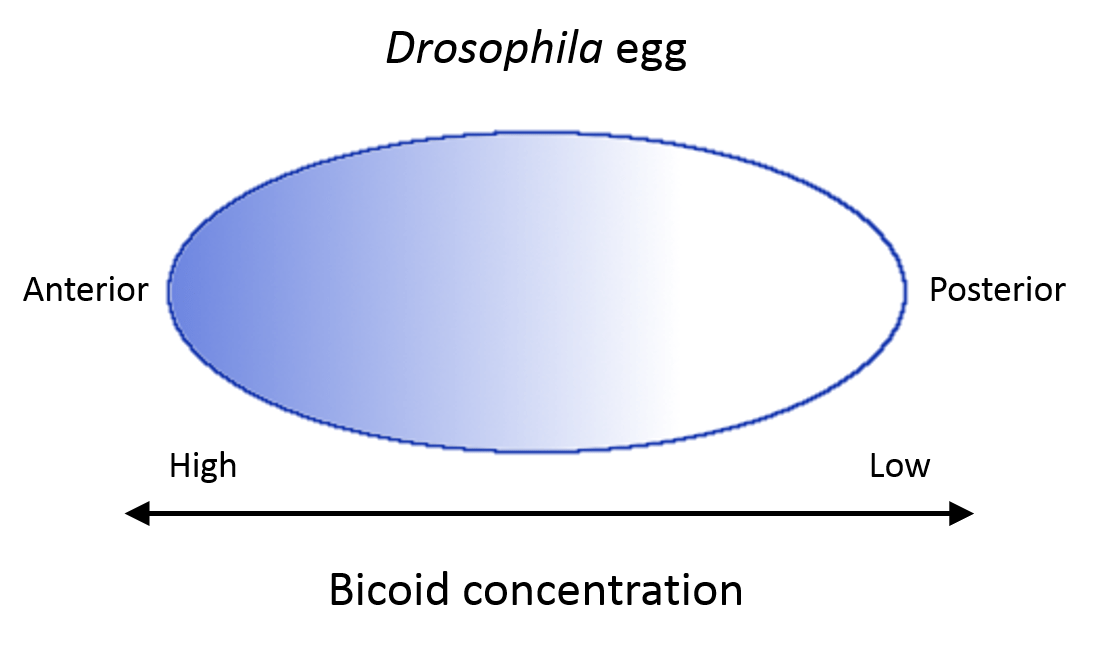
- Cytoplasmic determinants can be either mRNAs or proteins that are found in the egg prior to fertilization (they come from mom’s genome, not the embryo). They are asymmetrically distributed, so that after the first cleavage division, the two different resulting cells end up with different intrinsic information, which will then lead to different cell fates. One of the best understood cytoplasmic determinants is a factor called bicoid, which is present in a concentration gradient across the unfertilized eggs of Drosophila (fruit flies). The region of the egg with the highest concentration of bicoid becomes the anterior (head) portion of the embryo, while the region with the lowest bicoid concentration becomes the posterior (tail) region of the embryo. This future cell identity of anterior vs posterior is set after the very first cleavage division, where one cell gets nearly all the bicoid and the other cell has almost none. Cytoplasmic determinants are a key feature of protostome development and some deuterostomes, but they are not present in mammalian embryos.
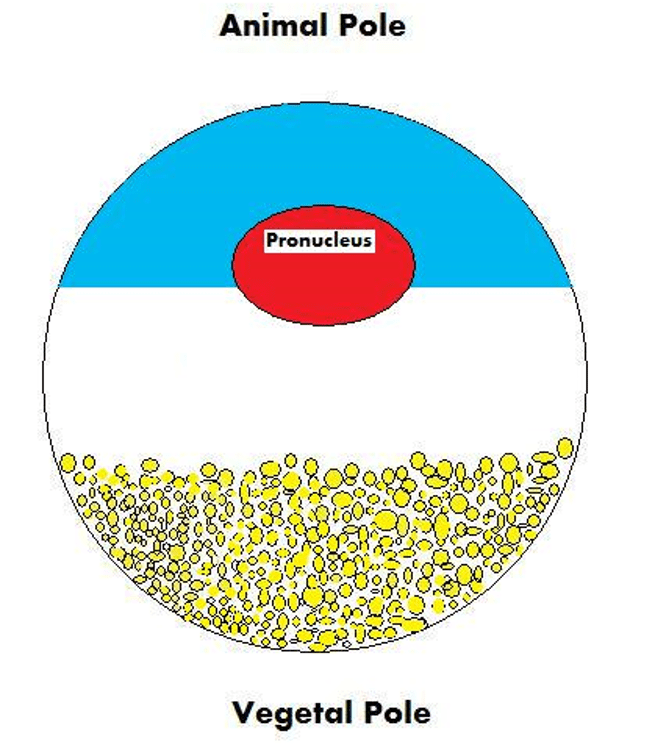
- Yolk polarity is a phenomenon that occurs in the eggs of animals which have large amounts of yolk in their eggs, such as amphibians, reptiles, birds, and many fish. Because the egg contains so much yolk, and the yolk is literally heavier than the rest of the egg, it sinks to the bottom of the egg due to gravity. The yolk is therefore asymmetrically distributed in the egg, and the region of the egg with very little yolk (also called the “animal pole”) will go on to become the anterior (head) structures of the embryo, while the region of the egg containing the heavier mass of yolk (also called the “vegetal pole”) will go on to become the posterior (tail) structures of the embryo.
- Induction, or cell-cell communication, is one of the five essential developmental processes, and occurs during development of all animal (and plant) species. Induction is a prime example of extrinsic information that leads to different cell fates among initially identical cells. Because mammalian embryos have no cytoplasmic determinants and have very small amounts of evenly-distributed yolk, induction is the primary process responsible for establishing the body axes in mammalian embryos.
The process of induction is important throughout development, and we will revisit it in the next reading on early animal development.
