
Learning Objectives
- Explain the significance, features, and consequences of gastrulation and organogenesis in early animal development
- List and describe adult tissue types in animals, and identify major organs arising from each embryonic germ layer, including: epidermal tissue, the nervous system, muscle and skeletal systems, circulatory system, reproductive organs, liver, pancreas, and epithelial lining of the digestive and respiratory systems.
- Recognize the roles and relationships of the four extra-embryonic membranes in amniotes (birds, reptiles, and mammals)
- Describe the roles of induction (cell-cell signaling) and regulation of gene expression in cell specialization and morphogenesis, using the notochord, the neural tube, and somites as examples
- Explain the relationship between Hox genes and segment identity in animals
Stages in early animal development
As we’ve already described, there are four general stages in early animal development:
- Fertilization: the process of a single sperm cell combining with a single egg cell to form a zygote.
- Cleavage: rapid, multiple rounds of mitotic cell division where the overall size of the embryo does not increase. The developing embryos is called a blastula following completion of cleavage.
- Gastrulation: the dramatic rearrangement (movement) of cells in the blastula to create the embryonic tissue layers. These tissue layers will go on to produce the tissues and organs of the adult animal.
- Organogenesis: the process of organ and tissue formation via cell division and differentiation.
Gastrulation and organogenesis together contribute to morphogenesis: the biological processes that result in an organism’s shape and body organization.
In this reading, we will discuss the last two steps above, gastrulation and organogenesis.
Development Step 3: Gastrulation
The information below was adapted from OpenStax Biology 43.6
At the end of cleavage, the typical blastula is a ball of cells with a hollow cavity in the middle (the blastocoel). The next stage in embryonic development is gastrulation, in which the cells in the blastula rearrange themselves to form three layers of cells and form the body plan. The embryo during this stage is called a gastrula. Gastrulation results in three important outcomes:
- The formation of the embryonic tissues, called germ layers. In organisms with three germ layers (triploblasts), the layers include the endoderm, ectoderm, and mesoderm (organisms with two germ layers – diploblasts – lack a mesoderm). Each germ layer will later differentiate into specialized tissues and organ systems.
- The formation of the embryonic gut, the archenteron.
- The appearance of the major body axes. Recall that in some species, the information specifying the body axes was already present during cleavage as a result of cytoplasmic determinants and/or yolk polarity, but the axes do not become visible until after gastrulation.
The specific details of gastrulation are different in among different animal lineages, but the general process includes dramatic movement of cells both across and inside the embryo:
In triploblasts (animals with three embryonic germ layers), one group of cells moves into the blastocoel, the interior of the embryo, through an invagination called the blastopore. These interior cells form the endoderm. Another group of cells move to completely surround the embryo, forming the ectoderm, and a third group of cells move into the locations in between the outer and inner layers of cells, to form the mesoderm. The endodermal cells continue through the interior of the embryo until they reach the other side, creating a continuous tract through the embryo; this tract is the archenteron, or embryonic gut. In protostomes, the blastopore becomes the embryo’s mouth; in deuterostomes, the blastopore becomes the embryo’s anus.
Diploblasts (animals with only two germ layers) do not have mesodermal cells. These animals, which include jellyfish and comb jellies, have radial rather than bilateral symmetry and have far fewer tissue types than triploblasts due to the lack of a mesoderm.
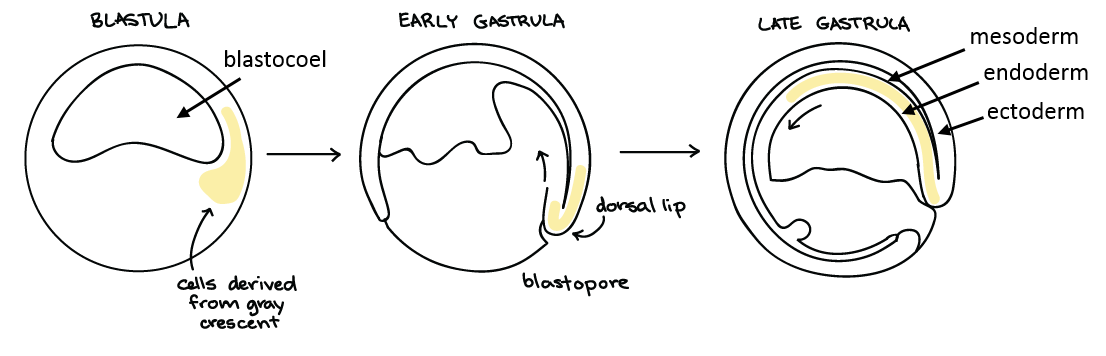
The three germ layers, shown below, are the endoderm, the ectoderm, and the mesoderm.
- The ectoderm gives rise to the nervous system the skin epidermis, and the epithelial lining of the mouth and rectum.
- The mesoderm gives rise to muscle cells and skeletal cells, the circulatory system, and the reproductive organs.
- The endoderm gives rise to many internal organs such as the liver and pancreas, and the epithelial lining of the digestive and respiratory systems.

This video provides an engaging overview of animal development, with a focus on gastrulation (and the fact that we’re all, like, tubes). Focus on the first 7:40 minutes:
And this video describes the different tissues and organs that arise from the different germ layers during human development:
From Germ Layers to Animal Tissues
The information below adapted from Khan Academy “Principles of Physiology”. All Khan Academy content is available for free at www.khanacademy.org
The cells in complex multicellular organisms are organized into tissues, groups of similar cells that work together on a specific task. Organs are structures made up of two or more tissues organized to carry out a particular function, and groups of organs with related functions make up the different organ systems.
Triploblasts have four primary adult tissues: epithelial tissue, connective tissue, muscle tissue, and nervous tissues, and these tissues arise from the three embryonic germ layers.
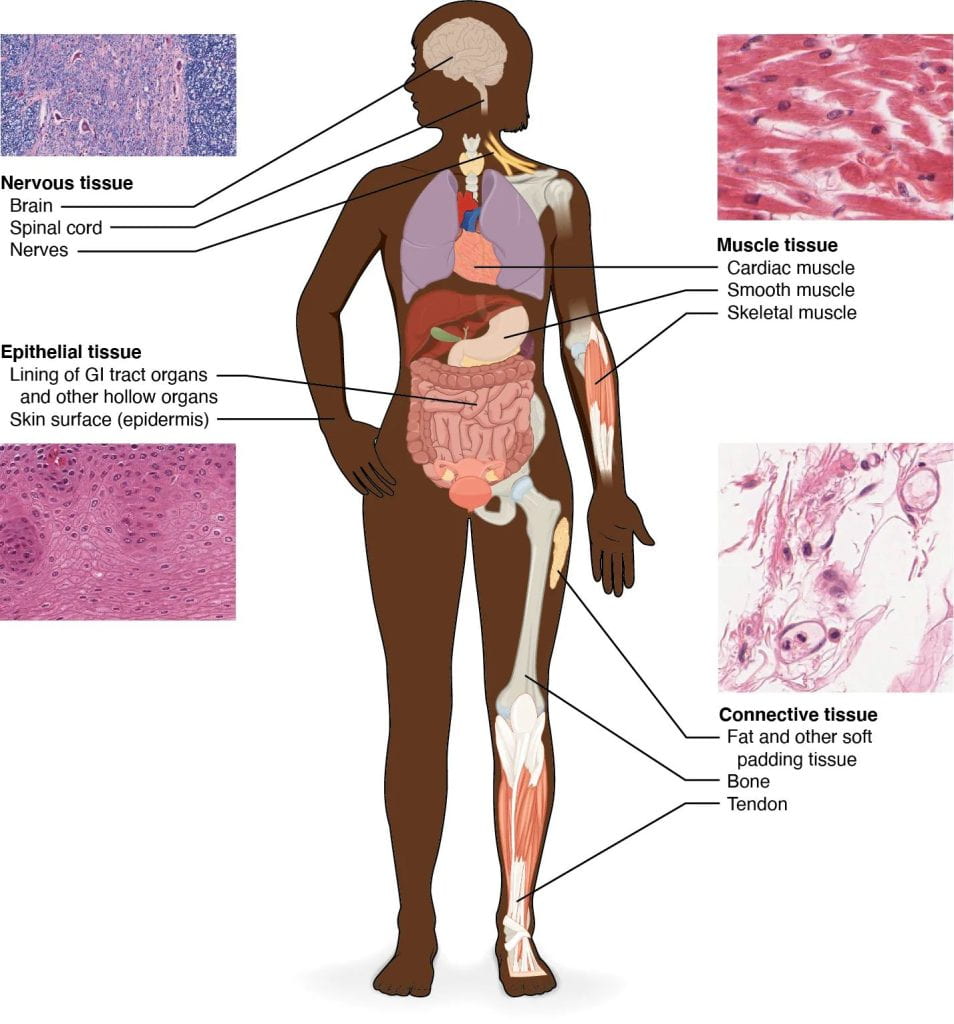
- Epithelial tissue consists of tightly packed sheets of cells that cover surfaces (such as the outside of the body) and line body cavities. For instance, the outer layer of your skin is an epithelial tissue, and so is the internal lining of your small intestine. The tight packing of epithelial cells lets them act as barriers to the movement of fluids and potentially harmful microbes. Epithelial cells are also “polarized”, meaning that they have a top and a bottom side. The apical (top) side of an epithelial cell faces the inside of a cavity or the outside of a structure and is usually exposed to fluid or air. The basal (bottom) side faces the underlying cells. For instance, the apical sides of intestinal cells have finger-like structures that increase surface area for absorbing nutrients.

- Connective tissue consists of cells suspended in some kind of extracellular matrix (“extra”cellular means “outside of” the cell). In most cases, the matrix is made up of protein fibers like collagen and fibrin in a solid, liquid, or jellylike ground substance. Connective tissue supports and connects other tissues.
- Loose connective tissue, shown below, is the most common type of connective tissue. It’s found throughout your body, and it supports organs and blood vessels and links epithelial tissues to the muscles underneath.
- Dense, or fibrous, connective tissue is found in tendons and ligaments, which connect muscles to bones and bones to each other, respectively.
- Other types of connective tissue includes adipose tissue (body fat), bone, cartilage, and blood.

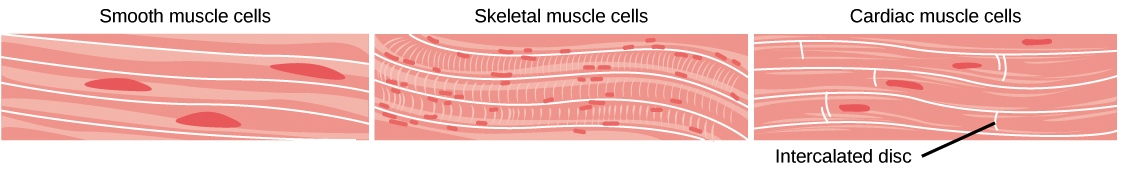
- Muscle tissue is essential for keeping the body upright, allowing it to move, and even pumping blood and pushing food through the digestive tract. Muscle cells, also called muscle fibers, contain the proteins actin and myosin, which allow them to contract. There are three main types of muscle: skeletal muscle, cardiac muscle, and smooth muscle.
- Skeletal muscle is what we commonly refer to as muscle in everyday life. Skeletal muscle is attached to bones by tendons, and it allows you to consciously control your movements.
- Cardiac muscle is found only in the walls of the heart. It’s not under voluntary control, so (thankfully!) you don’t need to think about making your heart beat.
- Smooth muscle is found in the walls of blood vessels, as well as in the walls of the digestive tract, the uterus, the urinary bladder, and various other internal structures. Smooth muscle is involuntary, not under conscious control. That means you don’t have to think about moving food through your digestive tract.
- Nervous tissue is involved in sensing stimuli (external or internal cues) and processing and transmitting information. It consists of two main types of cells: neurons and glia. The neurons are the basic functional unit of the nervous system: they generate electrical signals called action potentials that allow the neurons to convey information very rapidly across long distances. The glia mainly act to support neuronal function.

The video below walks through comparative animal anatomy and describes the four types of animal tissue:
The Four Extra-Embryonic Tissues in Amniotes
The information below was adapted from OpenStax Biology 29.4
The terrestrially-adapted amniotic egg is the defining characteristic of amniotes (reptiles, birds, and mammals). The evolution of amniotic membranes meant that the embryos of amniotes had their own aquatic environment built into their eggs, allowing them to exploit drier environments because they were less dependent on water for early development. This reduced dependence on water distinguished them from amphibians, which are restricted to moist environments due to their shell-less, non-amniotic eggs.
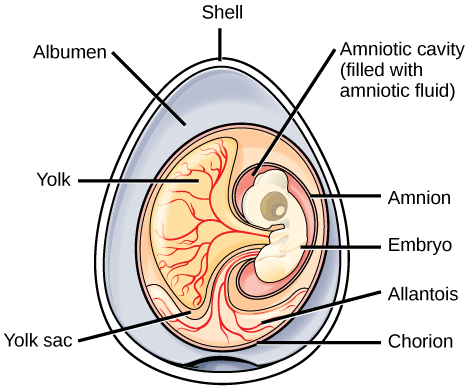
In amniotes that lay eggs (birds, most reptiles, and even some mammals), the shell of the egg provides protection for the developing embryo while allowing exchange of carbon dioxide and oxygen. The albumin, or egg white, provides the embryo with water and protein, and the egg yolk is the energy supply for the embryo (yolk is present in many other animals, such as amphibians). The eggs of amniotes also contain four additional extra-embryonic tissues: the amnion, chorion, allantois, and yolk sac, shown below. Extra-embryonic membranes are produced from the embryo, but they are not actually part of embryo (thus “extra”-embryonic). These extra-embryonic tissues are essential to the function of the amniotic egg:
- The amnion, or inner amniotic membrane, surrounds the embryo itself, enclosing the aqueous environment that the embryo develops in to protect the embryo from mechanical shock and support hydration.
- The chorion, which surrounds the embryo and yolk sac, facilitates exchange of oxygen and carbon dioxide between the embryo and the egg’s external environment.
- The allantois stores nitrogenous wastes produced by the embryo and also facilitates respiration in combination with the chorion.
- The yolk sac encloses the nutrient-rich yolk and transports nutrients from the yolk to the embryo (note the yolk sac is not the yolk itself, but is the membrane that surrounds the yolk)
Most mammals do not lay eggs (though some do!), but they still produce amniotic tissues that function as part of the placenta and umbilical cord, as shown below. Pregnancy in placental mammals is the result of internalization and incorporation of the amniotic egg into the uterus, resulting in direct nourishment embryo inside of the amniotic egg rather than laying it outside of the body with a predefined amount of yolk.
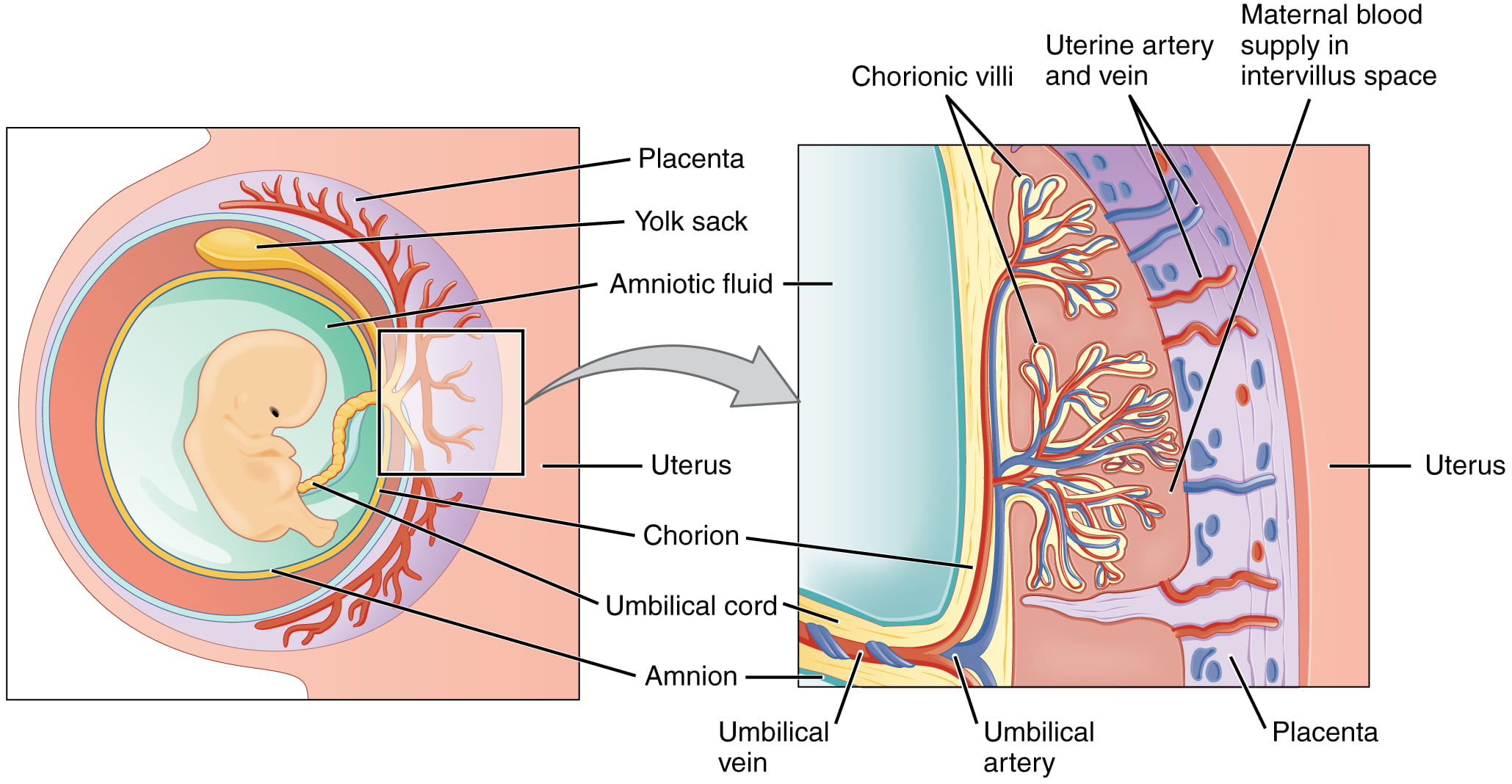
As you can see above, the chorion separates the fetal and maternal sides of the placenta, and the amnion surrounds the developing fetus. Just as in the amniotic egg:
- the amnion encloses the fluid-filled cavity to provide an aqueous environment for the developing fetus
- the chorion regulates gas exchange
- the allantois (not labeled), which functions in waste disposal, is part of the mammalian umbilical cord
- the yolk sac, consisting of blood vessels that transport nutrients to the embryo, is also part of the mammalian umbilical cord
Development Step 4: Organogenesis
The information below was adapted from OpenStax Biology 43.7
Gastrulation leads to the formation of the three germ layers; the germ layers ultimately lead to development of the different organs in the animal body. This process is called organogenesis.
In vertebrates, one of the primary steps during organogenesis is the formation of the nervous system. Interestingly, the nervous system originates from ectodermal, not mesodermal tissue. During the formation of the neural system, induction causes some cells at the edge of the ectoderm to become epidermis cells. The remaining cells in the center form the neural plate, which will go on to form the nervous system.
Immediately beneath the neural plate is a rod-shaped mesodermal structure called the notochord. The notochord signals the neural plate cells to fold over to form a tube called the neural tube, as illustrated below. During later development, the notochord will disappear (it goes on to form part of the spongy discs between the vertebrae), and the neural tube will give rise to the brain and the spinal cord.
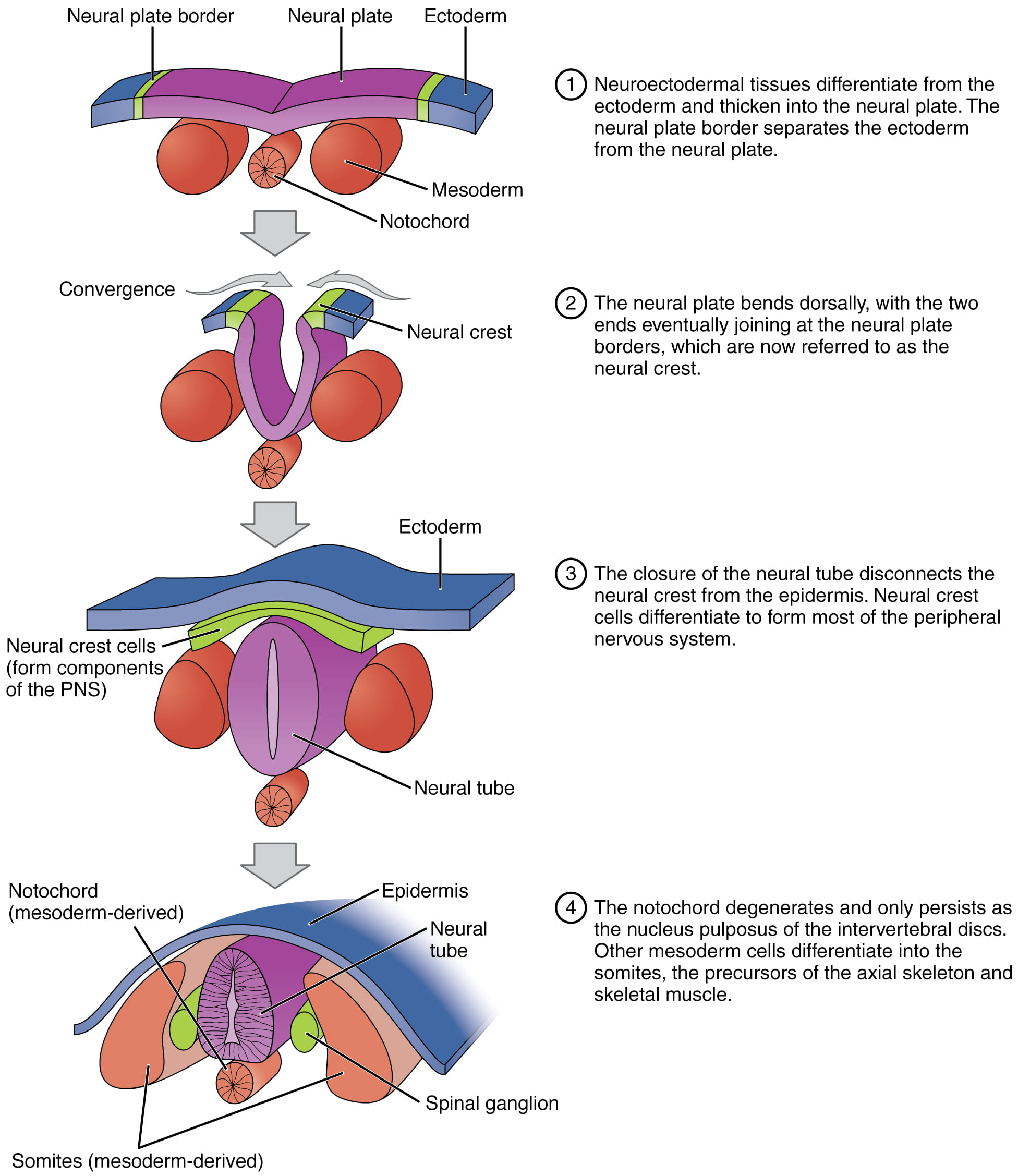
This video describes signaling from the notochord that results in neural tube formation:
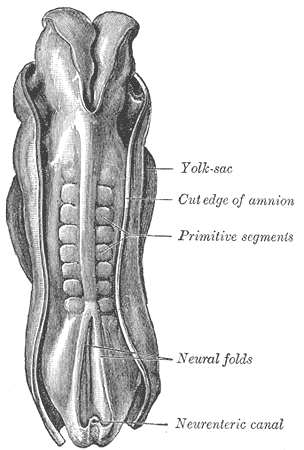
The mesoderm that lies on either side of the vertebrate neural tube forms a set of temporary structures called somites, shown below. Later in development, the cells within the somites will migrate to different parts of the body to develop into bone, skeletal muscle, and connective tissue of the skin. The specific pattern of induction from nearby tissues, including the ectoderm, the neural tube, the notochord, and surrounding mesoderm, will determine what type of tissue a particular region of a somite will become.
Hox Genes, Differential Gene Expression, and Segment Identity
Organogenesis (and development in general!) is characterized by changes in which specific genes are expressed in different cells. This differential gene expression, or turning on and turning off different genes, is what determines a specific cell’s form and function. It is also the process underlying differentiation (for more on this topic, see the Gene Regulation page in the Biological Principles textbook). For example, during differentiation, some cells in the ectoderm will express the genes specific to skin cells. As a result, these cells will differentiate into epidermal cells. Other ectoderm cells will move into the interior of the embryo to form the central nervous system, where they will express genes specific to the nervous system. The process of differentiation is largely regulated by induction, or cell-cell communication during development. How do induction and differential gene regulation work together to induce development of specific organs and body structure?
The information below was adapted from OpenStax Biology 27.1 and Khan Academy “Homeotic Genes.” All Khan Academy content is available for free at www.khanacademy.org
Changes in gene regulation during development are carefully regulated in both time and space. As we previously discussed, the eggs of protostomes and some deuterostomes contain cytoplasmic determinants, which cause cells of the developing embryo to have different identities as early as the first cell division (during cleavage). Cytoplasmic determinants are often regulatory genes that direct the expression of other genes, thus initiating a developmental “cascade” of changes in gene expression that ultimately lead to proper development of the animal. Each regulatory gene activates a new set of regulatory genes in the next set of cell divisions as the embryo progresses through development, as shown below.
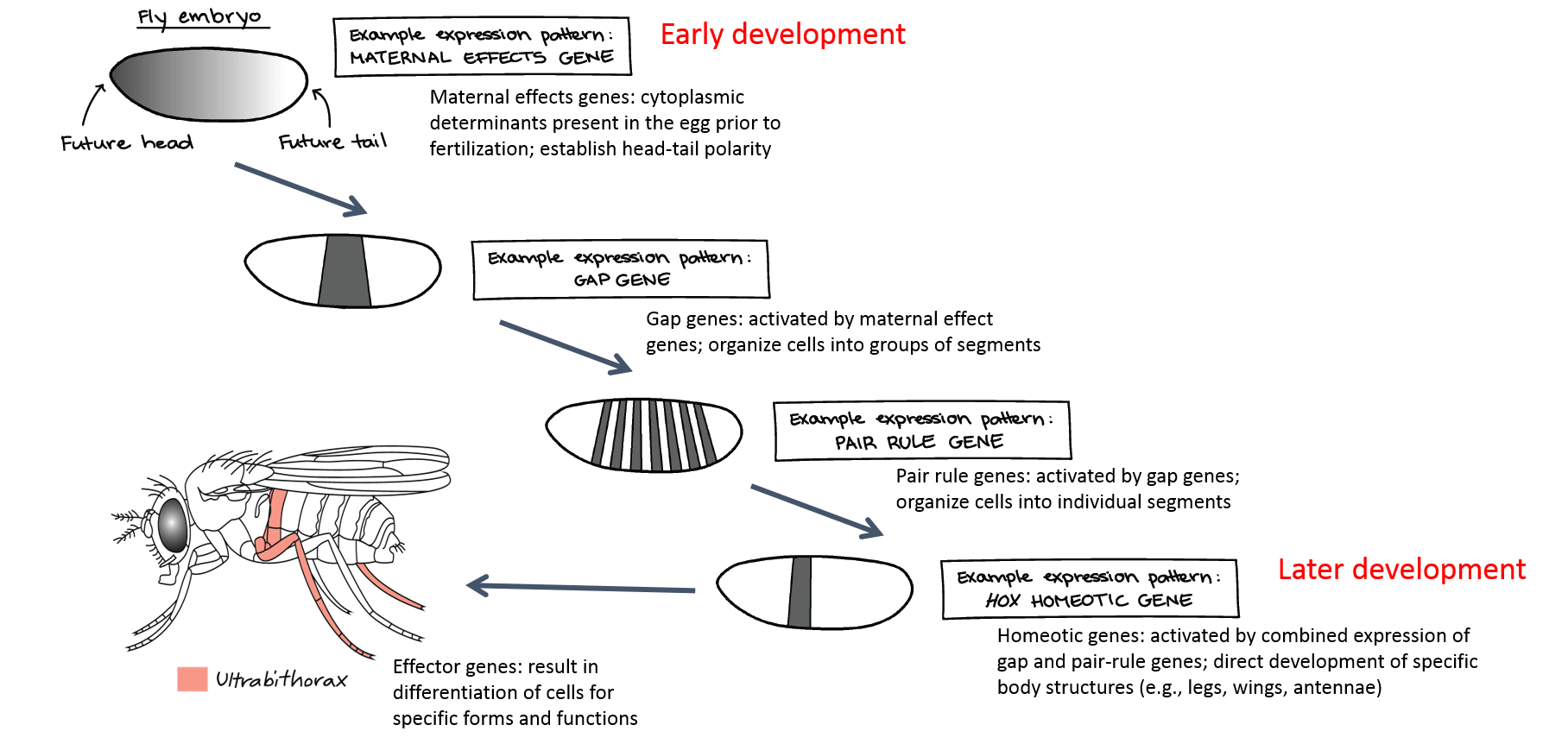
A key set of genes involved in differential gene expression and morphogenesis in animals are the homeobox or Hox genes. This family of genes is responsible for determining the general body plan, such as the number of body segments of an animal, the number and placement of appendages, and animal head-tail directionality. All animal phyla except sponges have a set of Hox genes. Each body segment is “specified” by a specific combination of Hox genes. In other words, Hox genes determine “segment identity,” or where along the body different body parts develop:

A single Hox mutation in the fruit fly can result in an extra pair of wings or even appendages growing from the wrong body part, as shown below.

While there are many genes that play roles in the morphological development of an animal, what makes Hox genes so powerful is that they serve as master control genes that can turn on or off large numbers of other genes. Hox genes do because they encode transcription factors that control the expression of numerous other genes. Hox genes are homologous in the animal kingdom, that is, the genetic sequences of Hox genes and their positions on chromosomes are remarkably similar across most animals because of their presence in a common ancestor, from worms to flies, mice, and humans. Hox genes have undergone multiple duplication events during animal evolution, with the additional genes allowing for more complex body types to evolve.

If a Hox 13 gene in a mouse was replaced with a Hox 1 gene, how might this alter animal development?
This video describes developmental regulatory genes in general and focuses on the importance of Hox genes in particular:
