Learning Objectives
- Compare and contrast the organization, structure, and function of gastrovascular cavities vs open and closed circulatory systems
- Compare and contrast the organization, structure, and function of vertebrate circulatory systems
- Differentiate between and describe the functions and structures of different types of blood vessels
- Recognize how blood pressure and blood velocity change across different categories of blood vessels
- Explain the processes of gas, nutrient, and fluid exchange between capillaries and tissues
- Identify and describe the functions of the different components of blood
Types of Animal Circulatory Systems
The information below was adapted from OpenStax Biology 40.1
Simple diffusion allows for exchange of water, nutrients, waste, and gases in animals that are only a few cell layers thick, but the circulatory system is the primary method used to transport nutrients and gases through most animal bodies. True circulatory systems share some common architecture, including:
- a circulatory fluid to facilitate nutrient and/or gas exchange
- tubes or vessels (arteries, capillaries, veins) that transport the circulatory fluid
- a muscular pump (heart) to move the circulatory fluid
Despite these common features, there is wide variation among animals in circulatory system structure:
- A gastrovascular cavity (previously discussed in the context of nutrient acquisition by animals) is not a “true” circulatory system because it lacks both a heart and blood vessels; however, it serves the role of a circulatory system because it is integral to the process of distributing the digested nutrients throughout the body.
- Gastrovascular cavities are common among diploblastic animals with radial symmetry, such as Ctenophora (comb jellies) and Cnidaria (coral, jellyfish, and sea anemones).
- Exchange of fluids is assisted by the pulsing of the animal’s body.
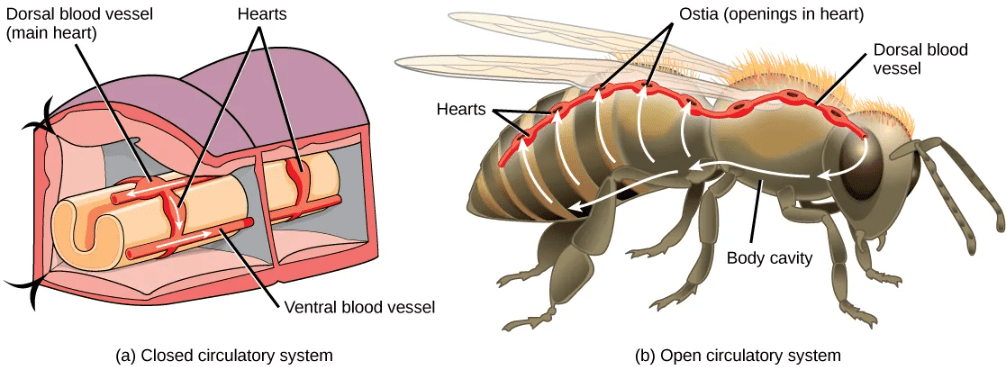
- An open circulatory system is a true circulatory system with circulatory fluid, vessels, and a muscular pump.
- Open circulatory systems are common (though not universal) among invertebrates, including arthropods and most mollusks.
- The circulatory fluid in open circulatory systems, called hemolymph, is pumped by the heart though vessels but then exits the vessels into a body cavity where it directly bathes the organs for nutrient and/or gas exchange. (Recall that insects do not use the circulatory system for gas exchange; this process is mediate by direct diffusion through the tracheae.
- Because the hemolymph is both pumped in vessels and also directly bathes the organs for nutrient exchange, the circulatory fluid is also the interstitial fluid in open circulatory systems.
- As the heart beats and the animal moves, the hemolymph circulates around the organs within the body cavity and then reenters the heart.
- The generally lower pressure required for an open circulatory system (compared to a closed circulatory system) translates to a lower energy cost for animals with this type of circulatory system.
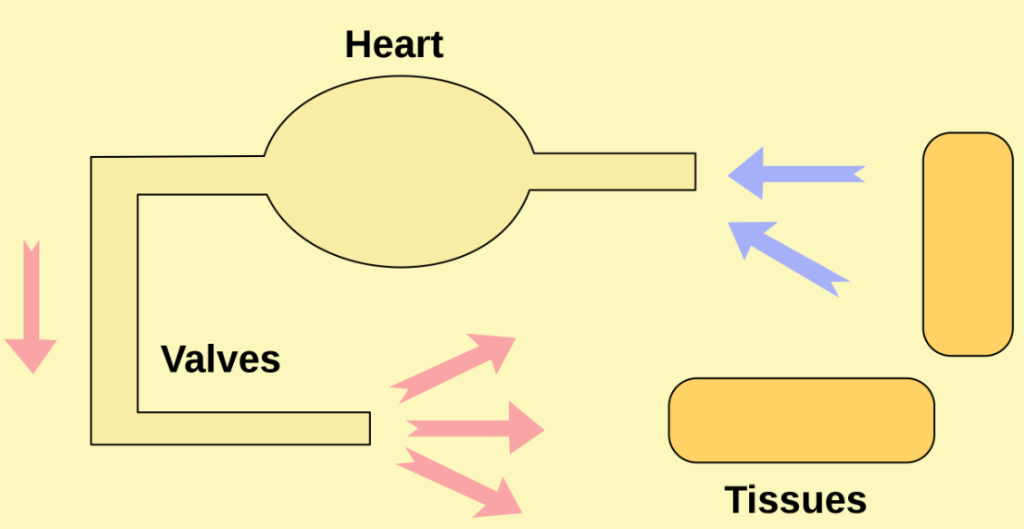
- A closed circulatory system is a true circulatory system where the circulatory fluid is pumped by a muscular heart and remains confined to vessels (the circulatory fluid is distinct from the interstitial fluid, unlike in open circulatory systems).
- Closed circulatory systems are universal among vertebrates and are common among many invertebrates including annelids (including earthworms) and cephalopods (including octopi and squids).
- Closed circulatory systems are under higher pressure than open systems, requiring a greater energy investment and subjecting the circulatory system to greater physical pressure and strain.
Variation Among Vertebrate Circulatory Systems
The information below was adapted from OpenStax Biology 40.1
All vertebrates have closed circulatory systems; however, there is wide variation in the structure and organization of closed circulatory systems among different vertebrate groups.
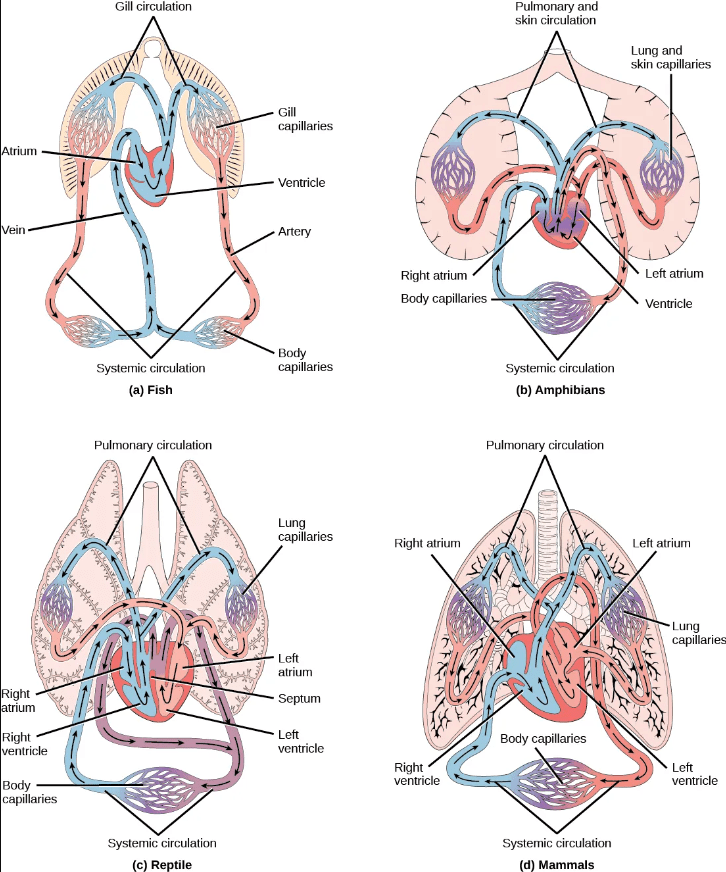
Two-chambered heart with a single circuit (single circulation): Fish have a single circuit for blood flow and a two-chambered heart that has only a single atrium (site of blood entry) and a single ventricle (site of blood exit):
- The atrium collects blood that has returned from the body, and the ventricle pumps the blood to the gills where gas exchange occurs and where the blood is re-oxygenated; this is called gill circulation.
- The blood then continues through the rest of the body before arriving back at the atrium; this is called systemic (body) circulation.
Three-chambered heart with two circuits (double circulation): Amphibians have a three-chambered heart that has two atria and one ventricle rather than the two-chambered heart of fish. As a result, blood flow is directed in two circuits: one through the lungs and back to the heart, which is called pulmonary circulation, and the other throughout the rest of the body and its organs including the brain (systemic circulation).
- The two atria receive blood from the two different circuits: the lungs/skin (where gas is exchanged with the environment) and the body. The blood then moves into the ventricle where some mixing of the blood occurs, though the mixing is mitigated by a ridge within the ventricle that diverts oxygen-rich blood through the systemic (body) circulatory system to deliver oxygen and deoxygenated blood to the pulmocutaneous (lungs + skin) circuit to acquire more oxygen and eliminate carbon dioxide. (Recall that in amphibians, gas exchange also occurs through the skin during pulmonary circulation in addition to the lungs.)
- Compared to a two-chambered heart, the advantage of the three chambered-heart is that the blood returns to the heart after becoming oxygenated for a second round of pumping into the body (systemic) circuit.
Most reptiles also have a three-chambered heart similar to the amphibian heart that directs blood to the pulmonary and systemic circuits. However, the ventricle is divided more effectively by a partial septum, which results in less mixing of oxygenated and deoxygenated blood.
- Some aquatic reptiles and turtles, as well as crocodilians, have a circulatory mechanism where the heart shunts blood from the lungs toward the stomach and other organs during long periods of submergence under water; this shunt allows the blood to bypass the pulmonary (lung) circuit when the animal is underwater and no gas exchange can occur, reducing the overall energy requirement for circulating blood throughout the body.
Four-chambered heart with two circuits (double circulation): In mammals and birds, the heart is divided completely into four chambers: two atria and two ventricles.
- Oxygenated blood is fully separated from deoxygenated blood, which improves the efficiency of double circulation and is probably required for supporting the warm-blooded lifestyle of mammals and birds.
- The four-chambered heart of birds and mammals evolved independently from a three-chambered heart. The independent evolution of the same or a similar biological trait is referred to as convergent evolution.
The image below shows simplified schematics comparing open and closed circulatory systems, including variations among the closed circulatory systems present in vertebrates:
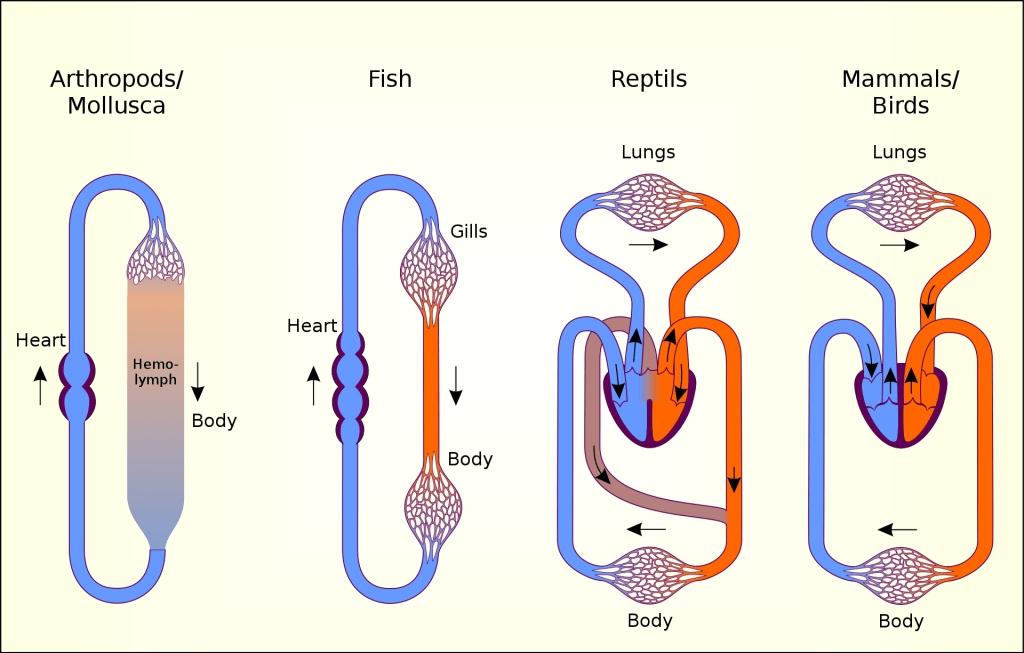
The image below shows more detailed diagrams comparing vertebrate circulatory systems:
The video below provides an evolutionary perspective on circulatory systems:
Types and Functions of Blood Vessels
The information below was adapted from OpenStax Biology 40.3
The blood from the heart is carried through the body by a complex network of blood vessels, each with distinct structures and functions:
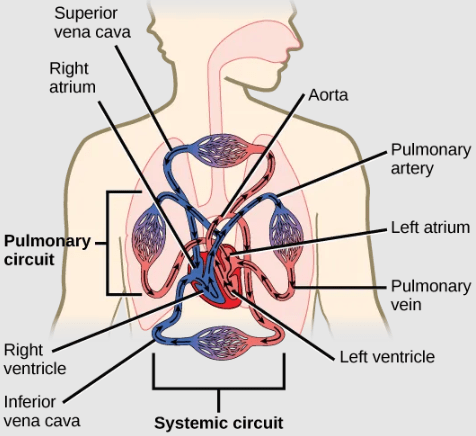
- Arteries take blood away from the heart, under high pressure. Arteries are defined by the fact that they carry blood away from the heart, not their oxygenation status: not all arteries carry oxygenated blood. Review the diagram below to identify arteries that carry deoxygenated blood. Arteries are lined with thick connective tissue and muscle tissue that support the artery wall and help to regulate the amount of pressure sustained by the artery. Arteries diverge into smaller arteries, which then diverge into smaller arteries called arterioles, to reach more deeply into the muscles and organs of the body. Arterioles diverge into capillaries.
- Capillaries are the sites for the exchange of nutrients, waste, and oxygen with tissues at the cellular level. Gas exchange in capillaries occurs both in the lungs, where capillaries acquire oxygen and release carbon dioxide, and also in the tissues where the opposite process occurs. Capillaries are very narrow, allowing for a single red blood cell to pass through at a time. Capillaries are also very thin, allowing for exchange of gas, nutrients, and fluid. Fluid that leaks out of capillaries can return back to the capillaries, and can also be brought back to the heart via the lymphatic system. The capillaries converge again into venules (small veins).
- Veins return blood to the heart. that connect to minor veins that finally connect to major veins that take blood high in carbon dioxide back to the heart. Veins are blood vessels that bring blood back to the heart, under low pressure. Veins are defined by the fact that they return blood to the heart: not all veins carry deoxygenated blood. Look at the diagram below to identify veins that carry oxygenated blood. Veins are lined with connective and muscle tissue, but these linings are thinner than in arteries. In addition, veins have valves to prevent the backflow of blood. Because veins have to work against gravity to get blood back to the heart, contraction of skeletal muscle assists with the flow of blood back to the heart.
This video describes the structure and function of different types of blood vessels:
Blood Pressure and Velocity Vary Throughout the Circulatory System
Blood exits the heart into arteries under high pressure and high velocity (speed). Blood pressure and velocity drop rapidly as the blood enters the capillaries, where speed slows to the slowest point in the circulatory system as individual red blood cells pass through the narrow capillary system. The blood velocity picks up in the veins, but pressure becomes even lower in veins.
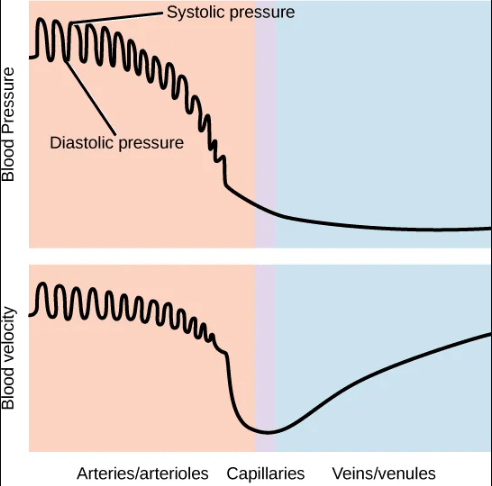
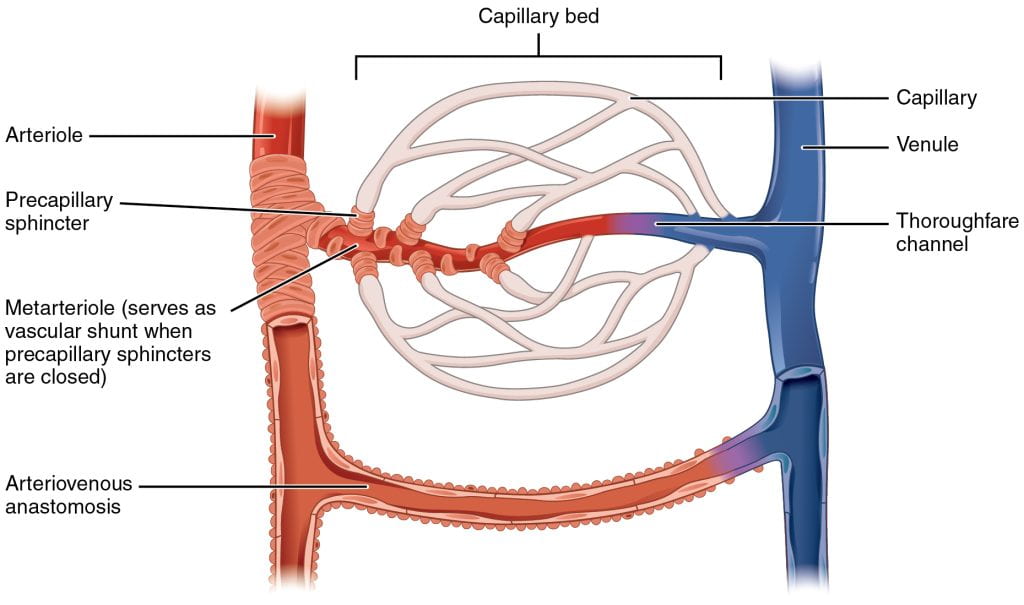
Blood flow and pressure can be regulated within the capillaries by the muscles lining the arterioles and venules on either side of the capillaries: vasoconstriction, narrowing of the blood vessels, increases blood pressure; vasodilation, widening of the blood vessels, reduces blood pressure.
Vasoconstriction and vasodilation are controlled by small muscles located at the starting point of each capillary bed: if the muscles are relaxed, then the blood will flow from the arteriole into the branches of the capillary blood. If all of the muscles are contracted, then the blood will bypass the capillary bed and go directly into the venule. These muscles allow precise control over when capillary beds receive blood flow; at any given moment, it is estimated that only about 5-10% of our capillary beds actually have blood flowing through them.
Gas, Nutrient, and Fluid Exchange Across Capillaries
The information below was adapted from OpenStax Biology 40.4
Exchange of gasses, nutrients, and fluid occurs only at the capillaries, which is facilitated by the slow rate that blood travels through the capillaries and the thin diffusion distance across the capillary walls. In addition to exchange of gas and nutrients, plasma (the watery connective tissue in blood), also leaves the capillaries and contributes to the interstitial fluid (the extracellular fluid surrounding the body tissues)
Most of the plasma-derived interstitial fluid returns to the capillaries before the capillaries converge into venules; this process occurs because of an osmotic and pressure gradient across the length of the capillary:
- Plasma leaves the capillaries due to relatively higher pressure at the arteriole end of the capillary.
- As plasma leaves the capillaries, the pressure in the interstitial fluid increases and the concentration of proteins and other large solutes remaining in the capillaries increases.
- The loss of the watery plasma creates a hyperosmotic solution within the capillaries, especially near the venules.
- As a result, most of the plasma diffuses back into the capillaries near the venules.
- Remaining plasma drains out from the interstitial fluid into nearby lymphatic vessels, where it passes through lymph nodes before it returns to the heart through the lymphatic system.
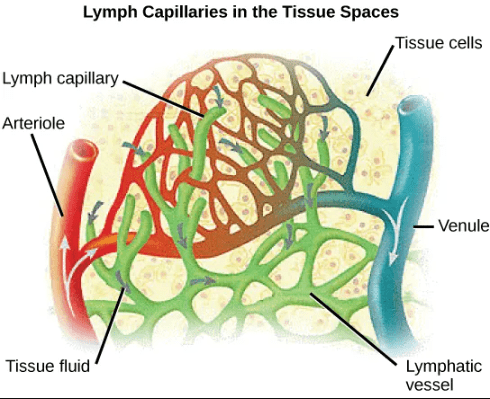
This video describes the function of the lymphatic system in conjunction with the circulatory system (stop at 5:40, when the discussion of immune function begins):
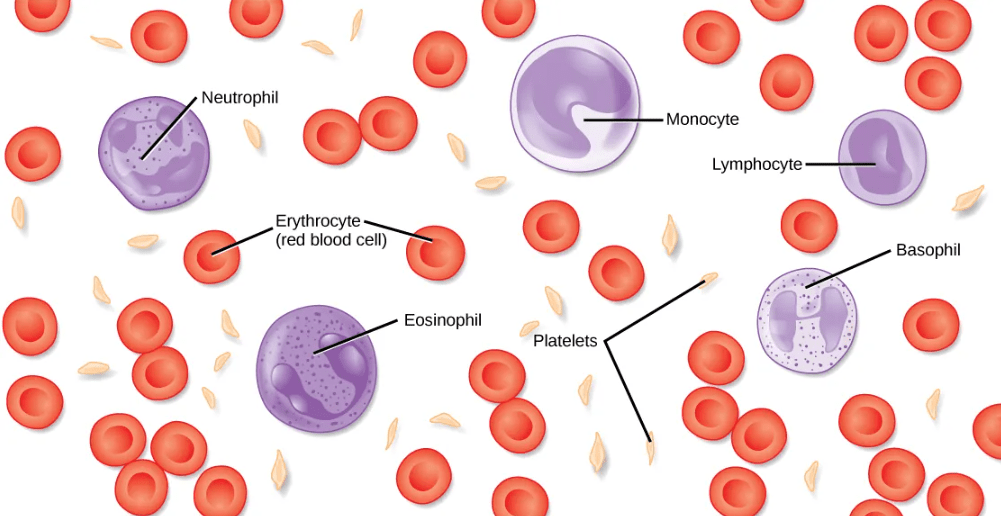
Function and Composition of Blood
The information below was adapted from OpenStax Biology 40.2
“Blood” is a term to describe the liquid that moves through the circulatory system, and it includes plasma (the liquid portion) and many different types of cells, each of which has distinct functions:
- plasma is the liquid portion of blood, which contains water, proteins, salts, lipids, and glucose
- red blood cells contain the protein hemoglobin, and these cells transport oxygen and carbon dioxide in the circulatory systems of vertebrates and many invertebrates.
- Invertebrates that utilize hemolymph rather than blood have alternatives to hemoglobin to bind and transport oxygen:
- Hemocyanin, a blue-green, copper-containing protein is found in mollusks, crustaceans, and some of the arthropods.
- Chlorocruorin, a green-colored, iron-containing pigment is found in four families of polychaete tubeworms.
- Hemerythrin, a red, iron-containing protein is found in some polychaete worms and annelids. Despite the name, hemerythrin does not contain a heme group and its oxygen-carrying capacity is poor compared to hemoglobin.
- white blood cells are primarily involved in the immune response to identify and target pathogens, such as invading bacteria, viruses, and other foreign organisms. White blood cells are formed continually; some only live for hours or days, but some live for years.
- platelets work with other factors to promote blood clotting at sites of tissue damage.