
Learning Objectives
- Place and identify the clade Animals on a phylogenetic tree within the domain Eukarya
- Identify and describe key traits and adaptations common to all animals, and those common to all Eumetazoans
- Identify, and describe key adaptations present among animal invertebrate lineages
- Differentiate between a select number of invertebrate groups using differences in their key adaptations
- Explain how the coevolution of insects and angiosperms led to adaptive radiations of both groups, and identify the geologic era in which this occurred
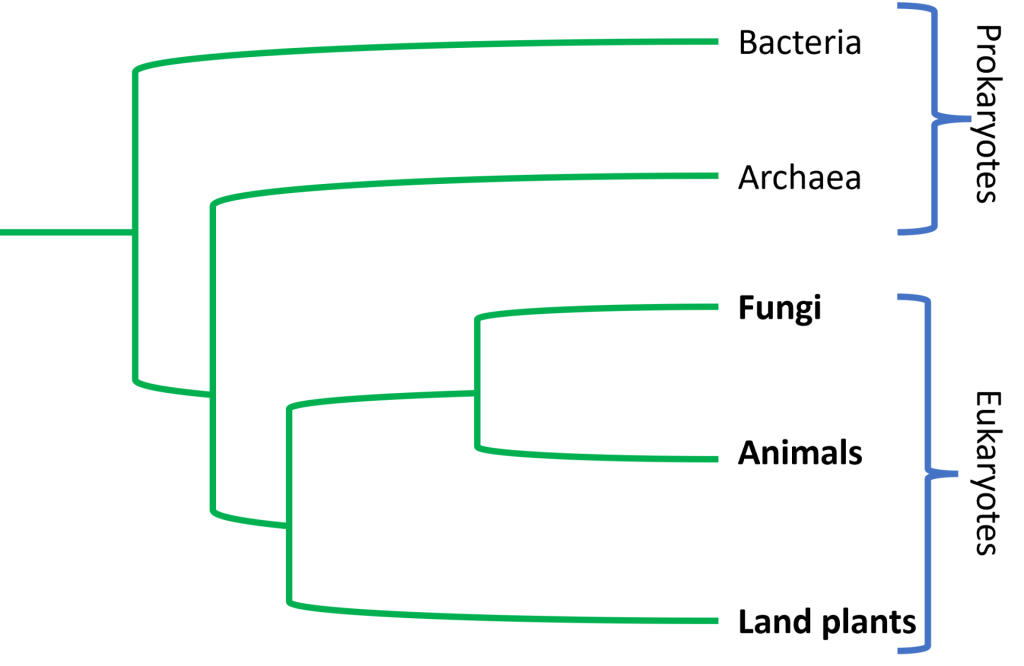
Animals on the Phylogenetic Tree of Life
Animals are a monophyletic group descended from an ancestral protist believed to be similar to modern day choanoflagellates. Fossil evidence suggests that multicellular animals first emerged around 800-900 million years ago in the Proterozoic Eon, and by the end of the Cambrian radiation(~542 MYA), animal life had began to greatly diversify. Today, although only 1.4 million species have been identified, there are somewhere between 8-5o million animal species estimated to be alive today.
Common Adaptations of Metazoans (Animals)
Animals are called metazoans (from Ancient Greek meaning “later animals”), and a subset of metazoans are categorized as eumetazoans (from Ancient Greek meaning “true animals”). Eumetazoa includes all animals except sponges (Porifera) and a group of amoeba-like “flat animals” (Placozoa). Eumetazoans possess true differentiated tissues, but sponges and placazoans do not have true tissues. The traits below are those shared by all animals (metazoans) and those with true differentiated tissues (eumetazoans).
- All Metazoa (all animals):
- Multicellular body plans
- Cells that lack cell walls
- Heterotrophy
- Movement at some point in their life cycle
- All Eumetazoa (all animals except Porifera and Placozoa):
- True differentiated tissues
- A nervous system (though not necessarily a central nervous system)
- Muscle cells
Key Evolutionary Adaptations among Invertebrate Lineages
What you might generally picture in your head as an “animal” may be a vertebrate species such as a dog, a bird, or a fish; however, concentrating on vertebrates gives us a rather biased and limited view of biodiversity because it ignores nearly 97(!) percent of all animals: the invertebrates. Invertebrate animals are those without a cranium (skull) or vertebral column. In addition to lacking vertebrae, most invertebrates also lack an endoskeleton and instead possess either a hydrostatic skeleton or an exoskeleton. A generally-accepted phylogeny of animals is below, but note that the evolutionary relationships among some of the of different groups of invertebrates are not yet clearly resolved.
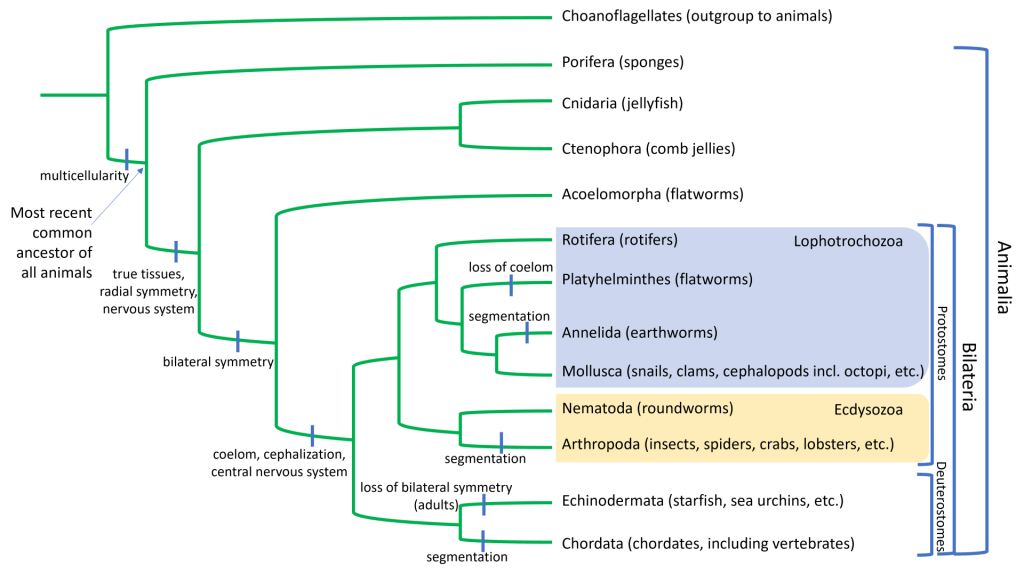
The first animals were invertebrates, and like the first fungi, the first animals were also aquatic. But unlike land plants, which experienced a single evolutionary transition to land, animals have experienced multiple water-to-land transitions in their evolutionary history. In other words: a terrestrial lifestyle is not a monophyletic trait. Here we will discuss some of the many adaptations among invertebrates, including those that facilitated the multiple water-to-land transitions in animal evolutionary history. Refer to the phylogenetic tree above to identify the major groups discussed with each evolutionary innovation described below.
The information below was adapted from CK-12 Biology for High School 11.2 Invertebrate Evolution and CK-12 Biology Advanced Concepts 15.3 Invertebrate Evolution – Advanced.

- Multicellularity and cell specialization: All animals are multicellular with specialized cell types, including Poriferans (sponges) and Placazoans, which lack true tissues but do have specialized cell types for digestion and reproduction.
- True tissues: All eumetazoans have true tissues: a group of similarly-structured cells that work together to carry out a specific function. The evolution of true tissues ultimately allowed for later evolution of complex organs and organ systems (an organ is a structure comprised of two or more specialized tissues that together carry out a specific function). The phylum Cnidaria (modern-day sea anemones, corals, jellyfish) likely includes what would have been the first eumetazoan.
- Tissue layers: Eumetazoans can be categorized by the number of embryonic tissue layers, groups of cells that form during early development and differentiate into specific tissues/organs:
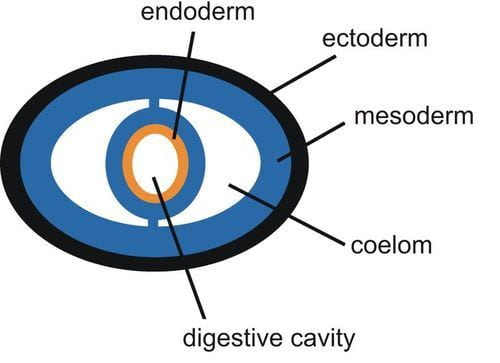
- Diploblasts: the first eumetazoan was a diploblast, meaning its tissues develop from two embryonic tissue layers, called the ectoderm (outer layer) and the endoderm (inner layer). Compared to organisms with three tissue layers (triploblasts), the presence of two tissue layers limits the number of tissue types that can be formed within an organism. Because diploblasts lack complex organ systems like respiratory systems, they are typically very thin or flat such that every cell in the body can exchange oxygen and carbon dioxide directly with the environment. Cnidarians and Ctenophores are diploblasts.
- Triploblasts and mesodermal tissue: The evolution of the mesoderm (middle layer) resulted in triploblasts, or animals with three embryonic tissue layers. The additional tissue layer, the mesoderm, differentiates into tissues such as muscle, complete digestive systems, and vascular systems. The evolution of these new tissue types allowed for more complex body structures and organ systems. The Acoelomorphs and all animals labelled Bilateria (see phylogeny above), are triploblastic.
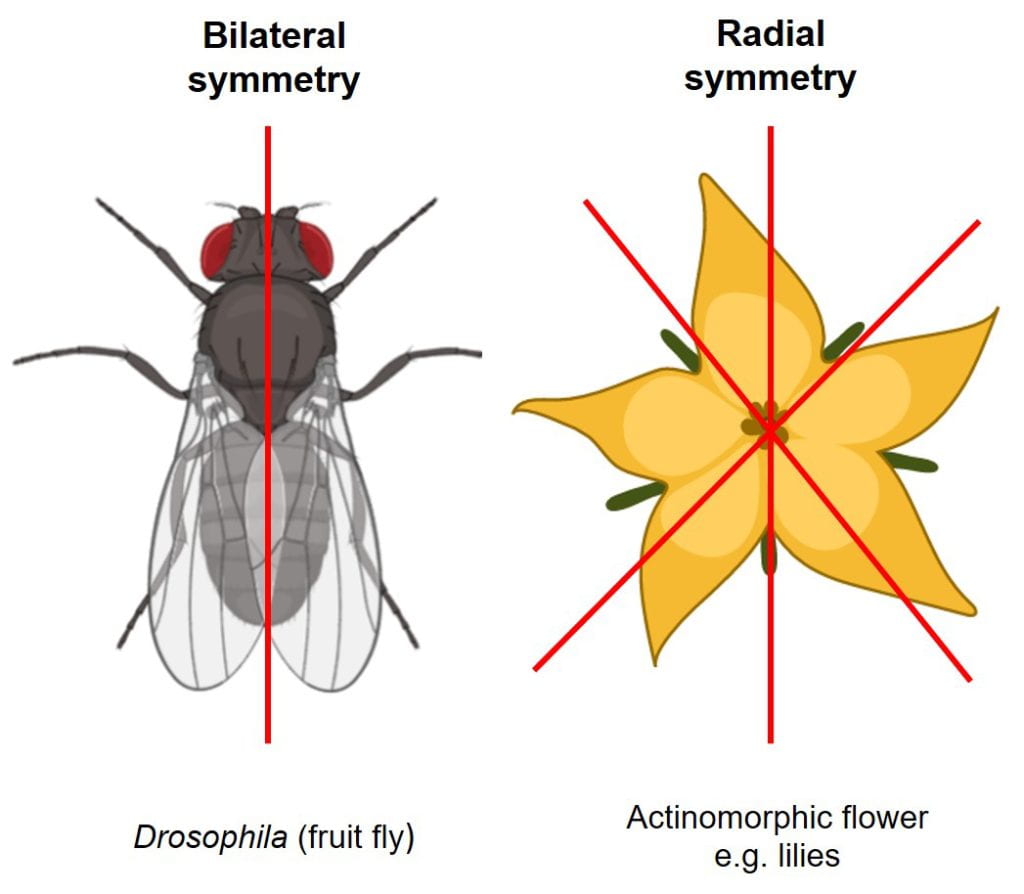
- Symmetry: Another trait that evolved early in animal evolution is symmetry (notice that sponges lack symmetry of any kind). Eumetazoans can also be categorized by the type of body symmetry present:
- Radial Symmetry: The first eumetazoans had radial symmetry, where the body has a repeating pattern around a central axis. Modern-day Cnidarians like jellyfish have radial symmetry. Radial symmetry is sufficient for slow-moving or sessile (non-moving) organisms but is less efficient at facilitating movement compared to bilateral symmetry.
- Bilateral symmetry and cephalization: Cephalization is the concentration of nerve tissue at one end of the body, forming a “head” region, and bilateral symmetry, or symmetry with a clear left-right or top-bottom, is a consequence of cephalization. Cephalization provides an evolutionary advantage in creating more controllable left-right or front-back movements. A simple form of cephalization + bilateral symmetry is a “worm-like” shape, and many invertebrates are worm-like, so it must be an advantageous body form! The earliest worm-like animals may have resembled modern-day Acoelomorpha, which are flatworm-like animals with simple body anatomy.
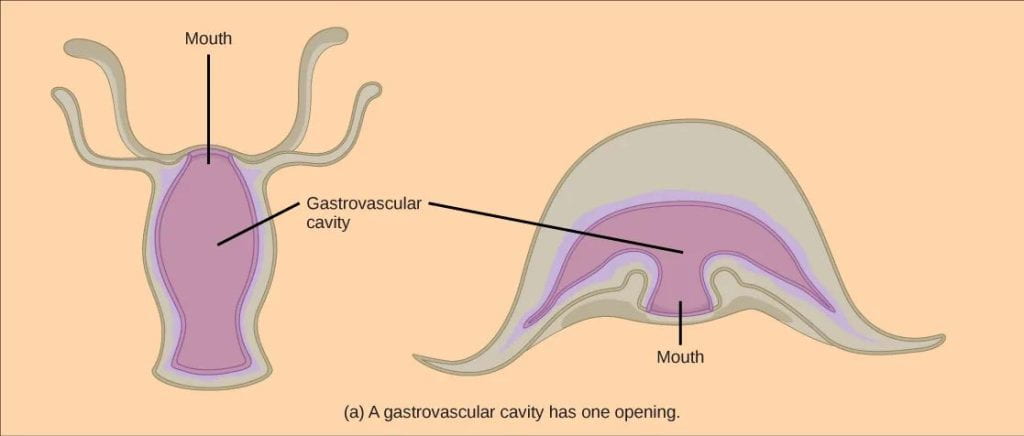
- Digestive systems: Animals are heterotrophic, which means they must obtain carbon from an organic source. Animals obtain carbon by eating another organism (for fungi, it generally means decomposing dead organic matter). Within their gut, an animal must digest, or break down the tissues of another organism into biologically-available molecules. Eumetazoans have two distinct types of digestives systems that can accomplish this task:
- Incomplete digestive system: The earliest eumetazoans possessed incomplete digestive system, where there is a digestive cavity (also called a gastrovascular cavity) with a single opening that serves as both mouth and anus. Incomplete digestive tracts are typically found in diploblastic organisms with radial symmetry, such as modern-day Cnidarians.
- Complete digestive system: Ancestors of Nematoda (roundworms) were also some of the first invertebrates to evolve a complete digestive system, with openings for both a mouth and an anus. The presence of separate openings allows for unidirectional movement of food and specialization of regions along the digestive tract. Complete digestive tracts are more efficient, allowing an animal to continue eating while digesting food and eliminating waste. Nearly all animals classified as Bilatera (see phylogeny above) have a complete digestive tract, with the exception of Platyhelminthes (flatworms) which lost this trait (and the coelom).
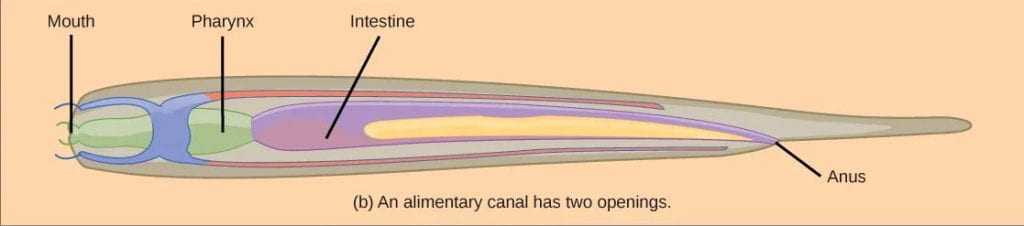
- Coelom (and pseudocoelom): A branch of early bilaterians evolved a structure called a pseudocoelom, or a partial body cavity filled with fluid. These animals were likely similar to modern roundworms (Nematoda), and this fluid-filled body cavity is the reasons roundworms are round, while flatworms – which lack a coelom – are flat. Further modification led to the development of a true coelom, which is lined on all sides by mesodermal tissue. Most animals classified as Bilateria have a coelom, with the exception of Platyhelminthes (flatworms) which lost this trait. A fluid-filled body cavity such as a coelom or pseudocoelom is evolutionarily significant for multiple reasons:
- the cavity allows room for internal organs to develop (often from mesodermal tissue) and remain attached to the mesodermal lining, which anchors them in place within the body
- the fluid in the cavity cushions and protects the internal organs from physical damage
- the pressure exerted by the fluid against the body walls gives the body internal support in the form of a hydrostatic skeleton; the hydrostatic skeleton in turn allows for efficient and coordinated movement driven by muscles
- Nervous systems: Evolution of neurons and nervous systems allowed for integration of sensory inputs and coordinated responses, such as movement toward or away from a stimulus. Except for Porifera (sponges) and Placozoa, all animals have some form of nervous system.
- Radially symmetric animals such as Cnidaria typically have a nerve net, a set of interconnected neurons without a brain or any sort of cephalization.
- The radially symmetric Echinodermata (sea stars, starfish, sea urchins), which evolved from bilaterally symmetric ancestors, have a central nerve ring around the mouth, and a set of radial nerves that extend out to the body.
- A central nervous system, consisting of a brain-like structure in the head region and at least one nerve cord that extends through the body, is present in most animals that demonstrate cephalization. Platyhelminthes have two nerve cords, Arthropoda have a ventral (belly) nerve cord, and Chordates (which includes a few lineages of invertebrates) have a dorsal (back) nerve cord.
- Respiratory systems and circulatory systems: Evolution of a respiratory system, with a dedicated structure for exchange of oxygen and carbon dioxide, allowed for evolution of more complex and large body sizes. A respiratory system also freed animals from needing a flattened body shape that would ensure all body cells could exchange oxygen and carbon dioxide with the environment. In many cases, a respiratory system is paired with a circulatory system* as a means to move gases and nutrients throughout the body. Interestingly, not all animals use a circulatory system to circulate oxygen. Insects have a series of air-filled tubes called tracheae that connect to the outside body to facilitate gas exchange.
- Segmentation: Segmentation has evolved multiple times in animal lineages, including in Annelida (earthworms), Arthropoda (insects, spiders, crabs, etc.), and Chordata (including invertebrate chordates). Segmentation is the division of the body into multiple subunits, which are often specialized into body parts specific to a region of the animal (e.g., segments of the head-thorax-abdomen regions of an insect). Segmentation increases flexibility and a wider range of motion by providing pivot points within the body.
- Limbs: Evolution of limbs has occurred multiple times in animal evolution, including in Arthropoda (insects, spiders, crabs, etc.) and Vertebrata (fish, birds, mammals, etc.). In both cases, limb evolution first appeared in aquatic species and ultimately facilitated the expansion of animals onto land.
- Wings: Wings have evolved multiple times in animal evolution, including in insects (Arthropoda) and birds and bats (Vertebrata).
The two videos below offer relatively humorous takes on the information above, and will help you connect these animal adaptations listed above to the different groups of invertebrate animals which possess them:
Exploring Traits among Major Invertebrate Groups
The information below was adapted from OpenStax Biology 28.1, OpenStax Biology 28.3, OpenStax Biology 28.4, OpenStax Biology 28.7
We will describe several major invertebrate groups below. The invertebrate phylogeny includes some unresolved and debated branching; however, the presence or absence of the adaptations listed above (and others) are used to distinguish groups of invertebrates. As you review this information, refer back to the phylogeny and list of adaptations above.
Porifera: Porifera is the phylum of sponges, which do not have true tissues, though they do have specialized cells. They also lack symmetry. The larval stages are motile, but adults are sessile (stationary).

Cnidaria: Cnidaria is a phylum that includes sea anemones, corals, and jellyfish. Cnidaria are radially symmetric diploblasts with true tissues and an incomplete digestive tract. Some species are motile, such as jellyfish, and others are non-motile as adults, such as sea anemones and corals. As they lack a respiratory system, their bodies and tissues are typically very thin to allow cells to exchange oxygen and carbon dioxide directly with the environment.

Acoelomorpha: Acoelomorpha is a subphylum of animals called “flatworms,” but they are distinct from true flatworms (Platyhelminthes, discussed below). Acoelomorpha are bilaterally symmetric but lack a gut and a coelom, as well as other organ systems that are present in most other bilaterally symmetric animals. Because they lack a respiratory system, their body organization must be flat to allow cells to exchange oxygen and carbon dioxide directly with the environment. Acoelomorpha may be descendants of the earliest lineage of animals that diverged from diploblasts, but they do not have all of the features of typical triploblasts. The location of Acoelomorphs on the animal phylogeny is still debated and unresolved.

Protostomes (Lophotrochozoa and Ecdysozoa): Protostomes are triploblastic animals where the mouth (as opposed to the anus) is the first part of the digestive tract to develop during embryogenesis. Protostomes are bilaterally symmetric with cephalization, and most also have a coelom, a complete digestive tract, and complex organ systems including a nervous system, circulatory system, respiratory system, and excretory system. Protostomes include two superphyla: Lophotrochozoa and Ecdysozoa.
Superphylum Lophotrochozoa: Lophotrochozoans are a diverse group of protostomes that include primarily soft-bodied invertebrates, and include two different phyla of “worms” (note that the term “worm” is not monophyletic). Lophotrochozoan phyla include Platyhelminthes, Annelida, Mollusca, Bryzoa, Rotifera, and others not described here.
- Platyhelminthes: Platyhelminthes is the phylum of “true” flatworms, and they are evolutionarily distinct from the Acoelomorpha. Platyhelminthes are triploblasts that lack a coelom, and they have an incomplete digestive tract. Similar to Acoelomorpha, their body plans are flat because they lack a respiratory system, so all cells must be in close proximity to the external environment to allow for gas exchange.

- Annelida: Annelids are a phylum of segmented worms that include earthworms. Annelids have a nervous system, a complete digestive tract, a circulatory system, and cutaneous respiration (they use the skin as their respiratory surface). Their muscles efficiently move a hydrostatic skeleton formed by the fluid-filled coelom. Their segmented body plan includes repeated morphological features in each body segment.

- Mollusca: Mollusca is the second-largest phylum of invertebrates (after Arthropoda). Molluscs are predominantly marine animals, soft-bodied animals including snails, slugs, nudibranchs (sea slugs), clams, mussels, oysters, squids, octopuses, cuttlefish, and nautiluses. They are morphologically diverse, but they all share the features that they are unsegmented and have a nervous system with two central nerve cords.

Superphylum Ecdysozoa, including Nematoda and Arthropoda: Ecdysozoans are protostomes named for the process of ecdysis, which refers to the shedding, or molting, of the exoskeleton. The phyla in this group have a hard cuticle that covers their bodies, which must be periodically shed and replaced in order for them to increase in size. The cuticle provides a tough but flexible exoskeleton that protects these animals from water loss, predators, and other aspects of the external environment.
- Nematoda: Nematodes are a phylum of animals called roundworms, because their overall morphology is cylindrical (in contrast to flat, like flatworms). Nematodes are triploblastic, bilaterally symmetric organisms with a pseudocoelom, or partial coelom. They are covered in an external cuticle that provides structural support, protection, and muscle attachment points for movement. They have a complete digestive system, a nervous system, and a hydrostatic skeleton combined with the cuticle-based exoskeleton. They lack a respiratory or circulatory system, which limits their overall size and diameter to allow for gas exchange by simple diffusion.
- Arthropoda: Arthropoda is the largest phylum in Animalia, containing an estimated 85% of all known living animal species. The most well-known features of arthropods are their hard exoskeletons made of chitin (the same composition as fungal cell walls), and their jointed appendages (which is what gives them their name). Arthropods are triploblastic, bilaterally symmetric organisms with a true coelom, segmentation, a complete digestive tract, a nervous system, respiratory systems, and excretory systems. Arthropods include five subphyla: Trilobitomorpha (trilobites, all extinct), Hexapoda (insects and relatives), Myriapoda (millipedes, centipedes, and relatives), Crustaceans (crabs, lobsters, crayfish, isopods, barnacles, and some zooplankton), and Chelicerata (horseshoe crabs, arachnids, scorpions, and spiders).

Deuterostomes: Echinoderms and Chordates: Deuterostomes are triploblastic animals where the anus (as opposed to the mouth) is the first part of the digestive tract to develop during embryogenesis. Deuterostomes include two phyla: Echinodermata and Chordata.
- Echinodermata: Echinoderms are a phylum of deuterostomes named for their spiny skin and include sea stars, sea urchins, sea cucumbers, sand dollars, and brittle stars. Echinoderms larvae are bilaterally symmetric, and adults have pentaradial (five-point radial) symmetry. They have a calcium-based endoskeleton, a nervous system consisting of a nerve ring and five radial nerves that extend outward along the arms, a complete digestive system, and a circulatory system without a heart. They have a unique “water vascular system” that functions in gas, nutrient, and water exchange, and also provides a mechanism for movement.

- Chordata: Chordates are the phylum of deuterostomes that consist of animals with a notochord (among other features listed below). If you know that vertebrates are members of phylum Chordata, then you may be wondering why Chordata is on this list since this reading is focused on invertebrates. In fact, Chordata includes invertebrates as well as vertebrates! Invertebrate chordates include tunicates (aka sea squirts) and lancelets. All chordates share a set of defining features at some point during their larval or adult stages: a notochord, a hollow dorsal nerve cord, pharyngeal slits, and a muscular post-anal tail.

Insect and Angiosperm Coevolution
Animal evolution begins during the Proterozoic Eon, with fossil evidence indicating multicellular animals were present prior to the Cambrian radiation. One pre-Cambrian fossil species, Kimberella, may have been an early Bilaterian. Animals then greatly diversified during the Cambrian radiation during the transition to the Phanerozoic Eon, ~542 MYA. The branches of the animal phylogeny were established by the end of the Cambrian period. These species were ocean-dwelling, with the first terrestrial animals (likely Arthropoda) appearing in the Ordovician period.

Insects first appear in the fossil record during the mid-Paleozoic Era. The fossil record indicates that the Carboniferous period of the late-Paleozoic featured the largest insects and other arthropods ever seen on Earth, including giant dragonflies with wingspans over 2 feet wide, giant scorpions over 2 feet long, and giant millipedes over 8 feet long. One of the major hypotheses to explain their large size is that higher atmospheric oxygen levels of the Carboniferous could support the metabolism of larger insects and other arthropods. Recall that insects use tracheae to deliver oxygen to internal cells; higher oxygen concentration would promote diffusion in longer tracheae. Oxygen comprised approximately 35% of the atmosphere during the Carboniferous period, compared to 21% today.
With the evolution of angiosperms (flowering plants) during the Mesozoic Era, insects and flowering plants developed mutualistic relationships based on pollination. During the Cenozoic Era, the subset of insects that act as pollinators experienced an adaptive radiation as flowering plants experienced their own adaptive radiation, with the two groups coevolving and influencing the traits of each other.
The video below describes the evolution of pollinating insects and associated coevolution of insects with flowering plants:

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}