Learning Objectives
- Recognize the relationship between meristems and indeterminant growth, and differentiate between primary and secondary growth
- Differentiate between apical and lateral meristems, and explain their different roles in plant growth
- Explain how the two lateral meristems contribute to secondary growth in woody stems
- Identify and classify the components of woody stems that are derived from secondary growth
Indeterminate Plant Growth: Meristems
The information below was adapted from OpenStax Biology 30.1
Animals exhibit determinant growth: they grow to a specific body size and shape and then stop growing, but plants exhibit indeterminate growth: they will continue adding new organs (leaves, stems, roots) as long as they have access to the necessary resources. Plants are able to continue growing indefinitely like this due to specialized tissues called meristems, which are regions of continuous cell division and growth. Meristematic tissue cells are essentially stem cells: they are undifferentiated, and they continue to produce cells that quickly differentiate, or specialize, and become permanent tissues (dermal, ground, and vascular).
Meristematic tissues consist of three types, based on their location in the plant:
- Apical meristems contain meristematic tissue located at the tips of stems and roots, which enable a plant to extend in length. Axillary buds also contain meristematic tissue.
- Lateral meristems facilitate growth in thickness or width in a maturing plant. Lateral meristems can also be called cambia (singular = cambium)
- Intercalary (also called basal) meristems occur only in some monocots, at the bases of leaf blades and at nodes (the areas where leaves attach to a stem). This tissue enables the monocot leaf blade to increase in length from the leaf base; for example, it allows lawn grass leaves to elongate even after repeated grazing or mowing.
Meristems contribute to both primary (taller/longer) and secondary (wider) growth:
- Primary growth is controlled by root apical meristems and shoot apical meristems.
- Secondary growth is controlled by the two lateral meristems, called the vascular cambium and the cork cambium. Not all plants exhibit secondary growth.
The video below provides a nice discussion of primary and secondary growth in plants (beginning at 2:20):
Primary Growth in Roots
The information below was adapted from OpenStax Biology 30.3
Root growth begins with seed germination. When the plant embryo emerges from the seed, the radicle of the embryo forms the root system. The tip of the root is protected by the root cap, a structure exclusive to roots and unlike any other plant structure. The root cap is continuously replaced because it is quickly damaged by the soil as the root pushes through soil. Behind the root cap, within the first centimeter or so, the root tip can be divided into three zones:
- The zone of cellular division is the location where new cells are being generated by mitosis. This process is driven by the activity of the apical meristem located immediately behind or under the root cap.
- The zone of cellular elongation is the location where the new cells are growing, or increasing in length, to add length to the root. This process requires uptake of water, which literally stretches the cells and increases their size.
- The zone of cellular maturation is the location where newly elongated cells complete their differentiation into the dermal, vascular, or ground tissues. Maturation is driven by changes in gene expression.

In another form of primary growth, some plants also have lateral roots that branch from the main tap root:
- The lateral roots originate from meristematic tissue in the pericycle, which is a layer of parenchyma cells (a type of ground tissue cell) that surrounds vascular cylinder in the center of the root (shown below).
- Once they have emerged, lateral roots then display their own primary growth by continually adding length to the lateral root. Note: lateral roots are NOT the same as secondary growth.


Primary Growth in Shoots
The information below was adapted from OpenStax Biology 30.2
Just as in roots, primary growth in stems is a result of rapidly dividing cells in the apical meristems at the shoot tip. Subsequent cell elongation then leads to primary growth. One difference is that there is no need for a structure like a root cap, because the stem does not have to force its way into the soil.
Most primary growth for many plants occurs primarily at the apical (top) bud, rather than the axillary buds (buds with meristematic tissue where the petiole joins the stem). This phenomenon is called apical dominance, where the apical bud prevents the growth of axillary buds that form along the branches and stems. Most coniferous trees exhibit strong apical dominance, thus producing the typical conical Christmas tree shape. If the apical bud is removed, then the axillary buds will start forming new stem growth. Gardeners take advantage of this process when they prune plants by cutting off the tops of branches, encouraging the axillary buds to grow out and give the plant a bushy shape.
Secondary Growth in Shoots (and Roots)
The process of secondary growth is controlled by the lateral meristems in both stems and roots. Lateral meristems include the vascular cambium and, in woody plants, the cork cambium (cambium is another term for meristem). Herbaceous (non-woody) plants mostly undergo primary growth, with very little secondary growth. Secondary growth, or wood, is noticeable in woody plants; it occurs in some eudicots, but occurs rarely in monocots.
The details below are specific to secondary growth in stems. While the principles are similar for secondary growth in roots, the details are somewhat different. We will discuss only the details specific to stems.
The vascular cambium contributes to the vast majority of secondary growth:
- The vascular cambium is located between the primary xylem and primary phloem within the vascular bundle; recall that vascular bundles are arranged in a ring near the periphery of eudicot stems, with the xylem located toward the interior and phloem toward the exterior of the bundle.
- The cells of the vascular cambium divide and form secondary xylem (tracheids and vessel elements) and secondary phloem (sieve elements and companion cells). These new cells appear between the primary xylem and primary phloem on either side of the cambium, with the secondary xylem toward the inside of the stem, and the secondary phloem toward the outside of the stem (see image below).
- The cells of the secondary xylem contain lignin, the primary component of wood, which provides strength and structural support.
- The xylem and the pith (ground tissue in the center of the stem) form the wood of a woody stem.
In woody plants, cork cambium forms an additional outermost lateral meristem:
- The cork cambium produces cork cells, which contain a waxy substance that can repel water.
- The phloem and the cork cells form the bark, which protects the plant against physical damage and helps reduce water loss.
- The cork cambium also produces a layer of cells known as phelloderm, which grows inward from the cork cambium. The cork cells, cork cambium, and phelloderm are collectively termed the periderm. The periderm substitutes for the epidermis in mature woody-stemmed plants.
The combined actions of the vascular and cork cambia result in secondary growth, with the activity of the vascular cambium contributing most of the increasing girth of the stem. These structures are illustrated below:

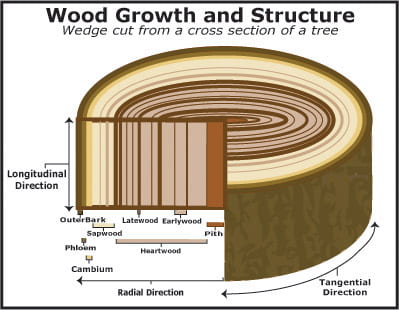
Components of Woody Stems and Contributions to Annual Tree Rings
A new layer of xylem and phloem are added by the vascular cambium each year during the growing season, creating new wood and new bark each year:
- Wood is composed of xylem and pith, the ground tissue found within the center of a woody stem; wood is everything from the vascular cambium toward the interior of the woody stem.
- Bark is composed of phloem and cork; bark is everything from the vascular cambium toward the exterior of the woody stem.
Over time, the wood gets larger in diameter, while the bark largely remains the same depth. Why is this this case?
- As the tree grows, it adds a new layer of xylem every year toward the inside of the ring of vascular cambium. As the tree becomes wider and wider, the interior xylem layers eventually fill with resin and become nonfunctional for carrying water, but they provide critical structural support as the tree becomes wider. This interior, nonfunctional xylem is called heartwood. The newer, functional xylem is called sapwood.
- A new layer of phloem is added each year toward the exterior of the ring of vascular cambium. The cork cambium, located outside of the vascular cambium, is also adding new layers of cork. The exterior-most layers of phloem become crushed against the cork cambium and are broken down within the bark. Thus a mature tree contains many interior layers of older, nonfunctional xylem deep within the stem, but only a small amount of older phloem.

This video describes the process and result of secondary growth in stems:
The activity of the vascular cambium results in annual growth rings. During the spring growing season, cells of the secondary xylem have a large internal diameter and their primary cell walls are relatively thin. This is known as early wood, or spring wood. During the fall season, the secondary xylem develops thickened cell walls, forming late wood, or autumn wood, which is denser than early wood. This alternation of early and late wood is due largely to a seasonal decrease in the number of vessel elements and a seasonal increase in the number of tracheids. It results in the formation of an annual ring, which can be seen as a circular ring in the cross section of the stem (shown below). An examination of the number of annual rings and their nature (such as their size and cell wall thickness) can reveal the age of the tree and the prevailing climatic conditions during each season.